All Areas of the Cerebral Cortex Share Common Histological Features
Different regions of the cerebral cortex have different functions. For example, the motor cortices (Chapter 10) project to the brainstem and spinal cord to initiate skilled, learned, conscious movement.
The occipital cortex processes visual information received from the retina of the eye (Chapter 14). The temporal cortex processes similar information from the ear (Chapter 17). However, even though different cortical regions have different functions, they have an underlying histological similarity. Therefore, cortical synaptic processing of information shares common features across regions, but differences in the origin of input signals and the destination of output signals contribute significantly to functional differences among regions. Cerebral cortical cells can work collectively over vast regions of the brain in such normal states as sleep and wakefulness and in such disease states as coma and seizures.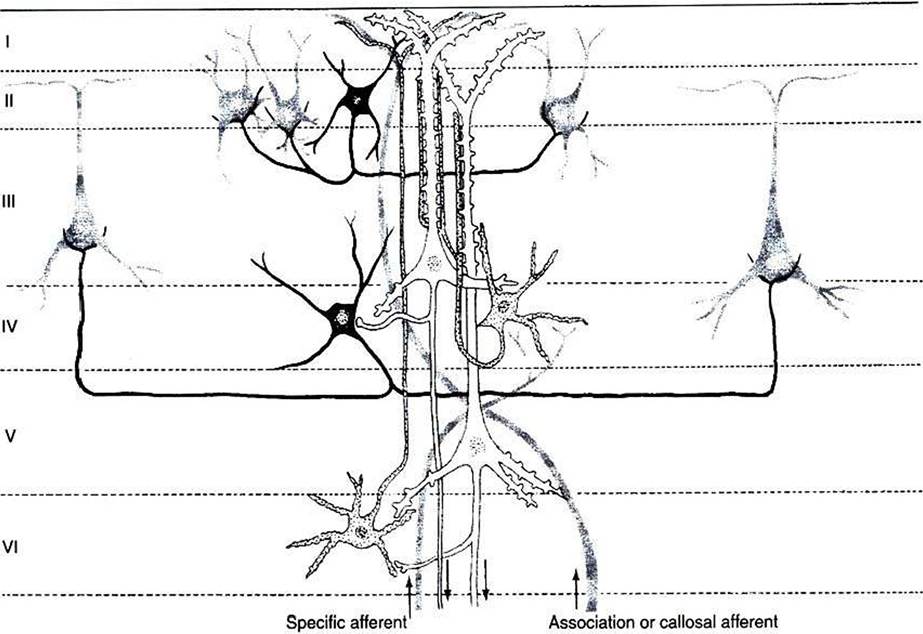
FIGURE 16-1 The principal neuron types and their interconnections have a basic similarity in the various regions of the cerebral cortex. Note that the two large pyramidal cells (white} in layers III and V receive multiple synaptic contacts from the star-shaped interneuron (stellate cell, stippled) in layer IV. Basket cell (black) inhibition is directed to the somata of cortical neurons. Major input to the cortex derives from specific thalamic nuclei (specific afferents) and is directed mostly to layer IV; association and callosal inputs (association and callosal afferents) have a more widespread termination pattern among the cortical layers. (From Kandel ER, Schwartz JH: Principles of neural science, ed 2, NewYork, 1985, Elsevier Science Publishing.)
The cerebral cortex contains several different cell types, but most belong to two major classes: pyramidal cells and stellate cells (Figure 16-1).
These cells are arranged in six layers (I-VI). The pyramidal cells, so named because their cell bodies are pyramid shaped, have dendrites projecting up toward the pial surface of the cortex, usually reaching and branching within layer I. These cells also have basal dendrites that extend horizontally from the cell body. Pyramidal cells are projection neurons, with their axons leaving their cortical region of origin and projecting to other parts of the central nervous system (CNS) or to a different region of cerebral cortex. Pyramidal cells are generally excitatory at their axon’s synapse. Stellate cells, so named because most have a starlike appearance, are local-circuit interneurons within the cortex and can be either excitatory or inhibitory. The majority of subcortical information arrives at the cortex through a massive input from the thalamic nuclei, most of which is targeted to layer IV. Input from some portions of the thalamus, as well as from other regions of cerebral cortex, have a more diffuse termination across cortical layers. Information arriving from cortical afferents is processed by local cortical circuitry, and pyramidal cells then carry the processed information to other CNS regions.As with other regions of the brain, the cerebral cortex contains about 10 to 50 times more glial cells than neurons. Three types of glial cells are present in the cortex: astrocytes, oligodendrocytes, and microglia. They do not develop action potentials, but as noted in Chapter 3, they may indirectly monitor neuronal electrical activity and modulate the effectiveness of neural communication. Glia also take up excess potassium ions, neurotransmitter, and toxins from the extracellular space and play a role in immune function. In addition, they help guide the course of developing neurons and stabilize the position of neurons, thus the origin of the term glia (“glue”).