Central Nervous System
Brain
The gross subdivisions of the adult brain include the cerebrum, cerebellum, and brainstem (Figs. 9-8 and 9-9). The cerebrum develops from the embryonic telencephalon. The components of the brainstem are defined in a number of ways; for our purposes, we include the diencephalon, midbrain, pons, and medulla oblongata as parts of the brainstem.
Telencephalon. The telencephalon, or cerebrum, comprises the two cerebral hemispheres, including the cerebral cortex, the basal nuclei, and other subcortical nuclei, and an aggregate of functionally related structures called the rhinencephalon. The telencephalon encloses the cavities of the lateral ventricles.
The surface area of the cerebrum in domestic mammals is increased by numerous foldings to form convex ridges, called gyri (singular gyrus), which are separated by furrows called fi ssures or sulci (singular sulcus). A particularly prominent fissure, the longitudinal fi ssure, lies on the median plane and separates the cerebrum into its right and left hemispheres. Unlike the spinal cord, in the cerebrum most of the neuronal cell bodies (i.e., the gray matter) are on the exterior. This layer of cerebral gray matter is called cerebral cortex. in humans and some animals, the cortical areas have been extensively mapped to localize specific sensory and
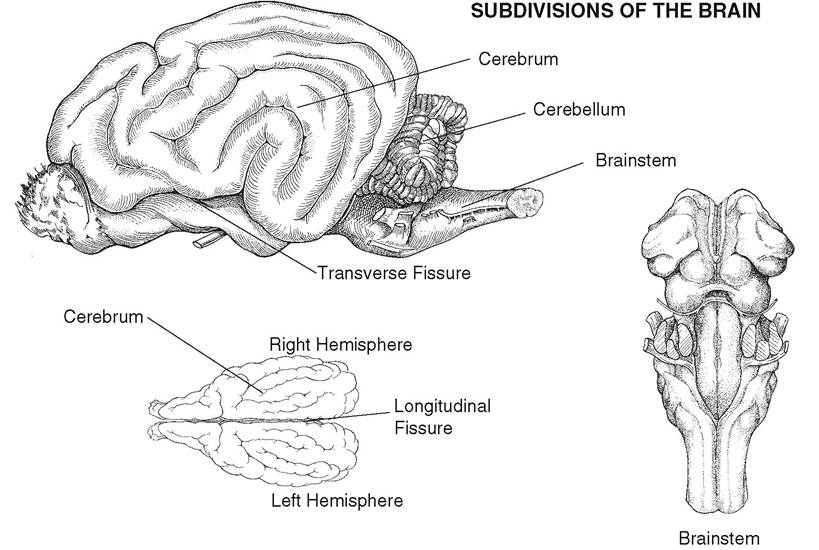
Figure 9-8. Subdivisions of the brain. The cerebrum consists of right and left cerebral hemispheres.
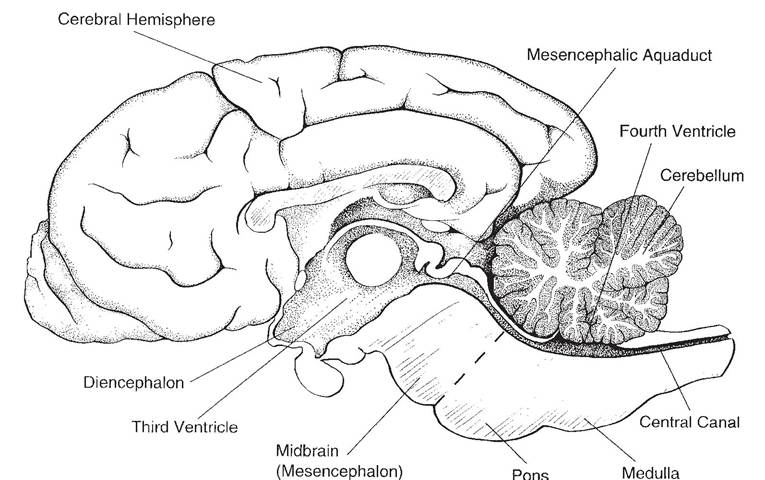
Figure 9-9. Midsagittal section of the brain. The lateral ventricles, one in each cerebral hemisphere, are not seen on this view, as they lie lateral to the midline.
motor functions. Anatomic regions defined by consistent gyri and sulci and general function are referred to as lobes.
The cerebral cortex is the site at which voluntary movements are initiated, sensations are brought to consciousness, and higher functions, such as reasoning and planning, take place.Deep to the cerebral cortex are aggregates of subcortical gray matter called the basal nuclei (an older term, basal ganglia, is discouraged, as the word ganglion usually refers to an accumulation of cell bodies outside the CNS). The basal nuclei are important in initiation and maintenance of normal motor activity. In humans, Huntington’s chorea and Parkinson’s disease are movement disorders caused by degeneration of parts of the basal nuclei.
The rhinencephalon is, from an evolutionary standpoint, one of the oldest parts of the cerebrum. It comprises a series of ventral and deep cortical structures associated primarily with the sense of smell (olfaction). The rhinencephalon has prominent connections to the parts of the brain that control autonomic functions, emotional behaviors, and memory, a fact that accounts for the striking ability of odors to affect these functions.
Diencephalon. The diencephalon is a derivative of the prosencephalon. The thalamus, epithalamus, hypothalamus, and the third ventricle are included in the diencephalon.
The thalamus is an important relay center for nerve fibers connecting the cerebral hemispheres to the brainstem, cerebellum, and spinal cord. The epithalamus, dorsal to the thalamus, includes the pineal gland, which is an endocrine organ in mammals. Its primary secretion, melatonin, appears to be important in circadian (daily) rhythms and sleep cycles. In addition, activity of the pineal gland is likely to be important in species with markedly seasonal reproductive cycles.
The hypothalamus, ventral to the thalamus, surrounds the ventral part of the third ventricle and comprises many nuclei that function in autonomic activities and behavior. Attached to the ventral part of the hypothalamus is the hypophysis, or pituitary gland, one of the most important endocrine glands.
The neuronal connections between the hypothalamus and the hypophysis constitute a critical point of integration of the two primary communication systems of the body, the nervous and endocrine systems.Mesencephalon. The mesencephalon, or midbrain, lies between the diencephalon ros- trally and the pons caudally. The two cerebral peduncles and four colliculi are the most prominent features of the midbrain.
The two cerebral peduncles, also called crura cerebri (singular crus cerebri), are large bundles of nerve fibers connecting the spinal cord and brainstem to the cerebral hemispheres. These peduncles consist largely (but not exclusively) of descending motor fiber tracts.
The colliculi (corpora quadrigemina) are four small bumps (colliculus is Latin for little hill) on the dorsal side of the midbrain. They consist of right and left rostral colliculi and right and left caudal colliculi. The rostral colliculi coordinate certain visual reflexes, and the caudal colliculi are relay nuclei for audition (hearing).
Metencephalon. The metencephalon includes the cerebellum dorsally and the pons ventrally. The cerebellum features two lateral hemispheres and a median ridge called the vermis because of its resemblance to a worm. The surface of the cerebellum consists of many laminae called folia. in the cerebellum, like the cerebrum, the white matter is central, and the gray matter is peripheral in the cerebellar cortex. The cerebellum is critical to the accurate timing and execution of movements; it acts to smooth and coordinate muscle activity.
The pons is ventral to the cerebellum, and its surface possesses visible transverse fibers that form a bridge from one hemisphere of the cerebellum to the other. Many other fiber tracts and cranial nerve nuclei make up the remainder of the pons.
Myelencephalon. The myelencephalon becomes the medulla oblongata in the adult. it is the cranial continuation of the spinal cord, from which it is arbitrarily distinguished at the foramen magnum.
The medulla oblongata (often simply called the medulla) contains important autonomic centers and nuclei for cranial nerves.Ventricular System. The ventricles of the brain develop from the lumen of the embryonic neural tube (Fig. 9-10). Right and left lateral ventricles lie within the respective cerebral hemispheres. They communicate with the midline third ventricle by way of two interventricular foramina. The third ventricle is surrounded by the diencephalon. it connects with the fourth ventricle by way of the narrow mesencephalic aqueduct (cerebral aqueduct) passing through the midbrain. The fourth ventricle, between the cerebellum above and pons and medulla below, communicates with the subarachnoid space surrounding the CNs through two lateral apertures.
Each ventricle features a choroid plexus, a tuft of capillaries that protrudes into the lumen of the ventricle. The plexus of capillaries is covered by a layer of ependymal cells that are continuous with the lining membrane of the ventricles.
The choroid plexus is responsible for forming the bulk of the cerebrospinal fluid (CSF) that fills the ventricular system and surrounds the CNs. A smaller contribution to that formation is made by the ependyma lining the ventricles. CSF is continuously formed, circulated, and reabsorbed in the spaces of the nervous system; its entire volume is replaced several times a day.
The circulation of cerebrospinal fluid begins in the two lateral ventricles (where the majority is produced). it flows through the interventricular foramina into the third ventricle, then by way of the cerebral aqueduct into the fourth ventricle, and finally through the lateral apertures into the subarachnoid space, where it surrounds both the brain and spinal cord. CSF is reabsorbed back into the venous blood via special modifications of the meninges called arachnoid granulations. These tiny structures protrude into the blood-filled dural sinuses (discussed later) and act as one-way valves for the return of CSF from the subarachnoid space to the venous blood.
Reabsorption of CSF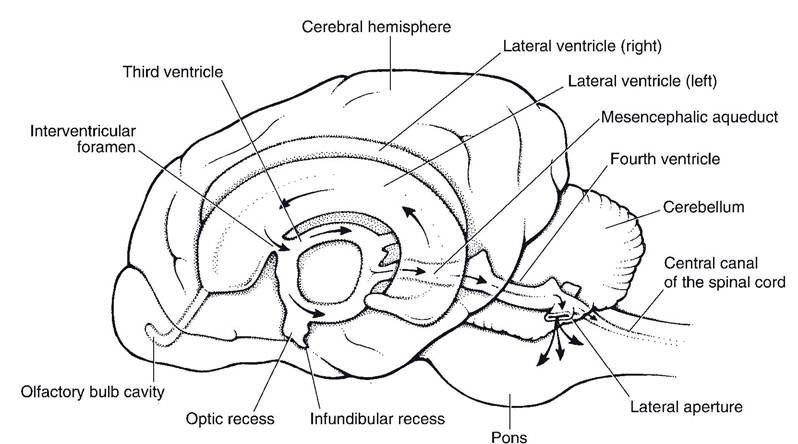
Figure 9-10. Ventricular system of the brain. Arrows indicate the direction of flow of cerebrospinal fluid (CSF), which is produced in the ventricles and flows via the lateral apertures into the subarachnoid space surrounding the brain and spinal cord. (Reprinted with permission of Wiley-Blackwell from Smith B.J. Canine Anatomy. Philadelphia: Lippincott Williams & Wilkins, 1999.)
is passive, driven by the pressure gradient across the arachnoid granulations (i.e., the higher pressure of CSF within the subarachnoid space drives the CSF into the low pressure sinus).
Any obstruction in cerebrospinal circulation can injure the brain, since the choroid plexuses produce CSF independent of the pressure within the ventricular system. Buildup of CSF will expand the affected ventricular spaces at the expense of nervous tissue, which can be markedly compressed as a result. If this occurs during development (as often happens when the mesencephalic aqueduct is inadequately formed), the head can become markedly enlarged and the cerebral tissue extremely compressed. This is the condition called hydrocephalus (literally, water head). In an older individual whose cranial bones are fully formed and fused at the sutures, buildup of CSF does not visibly enlarge the head; the elevated pressure inside the cranial vault, however, can profoundly affect the brain within.
Meninges
The connective tissue coverings of the brain and spinal cord are the meninges (singular meninx). They include, from deep to superficial, the pia mater, the arachnoid, and the dura mater (Fig. 9-11).
The pia mater, the deepest of the meninges, is a delicate membrane that invests the brain and spinal cord, following the grooves and depressions closely. The pia mater forms a sheath around the blood vessels and follows them into the substance of the CNS.
The middle meninx arises embryologically from the same layer as the pia mater but separates from it during development so that a space forms between them.
Remnants of their former connection in the adult take the form of many filaments of connective tissue that extend between them. Because of the weblike appearance of these filaments, this middle layer is called the arachnoid (arachnoidea, arachnoid mater), and the connecting filaments are the arachnoid trabeculae. Together, the pia mater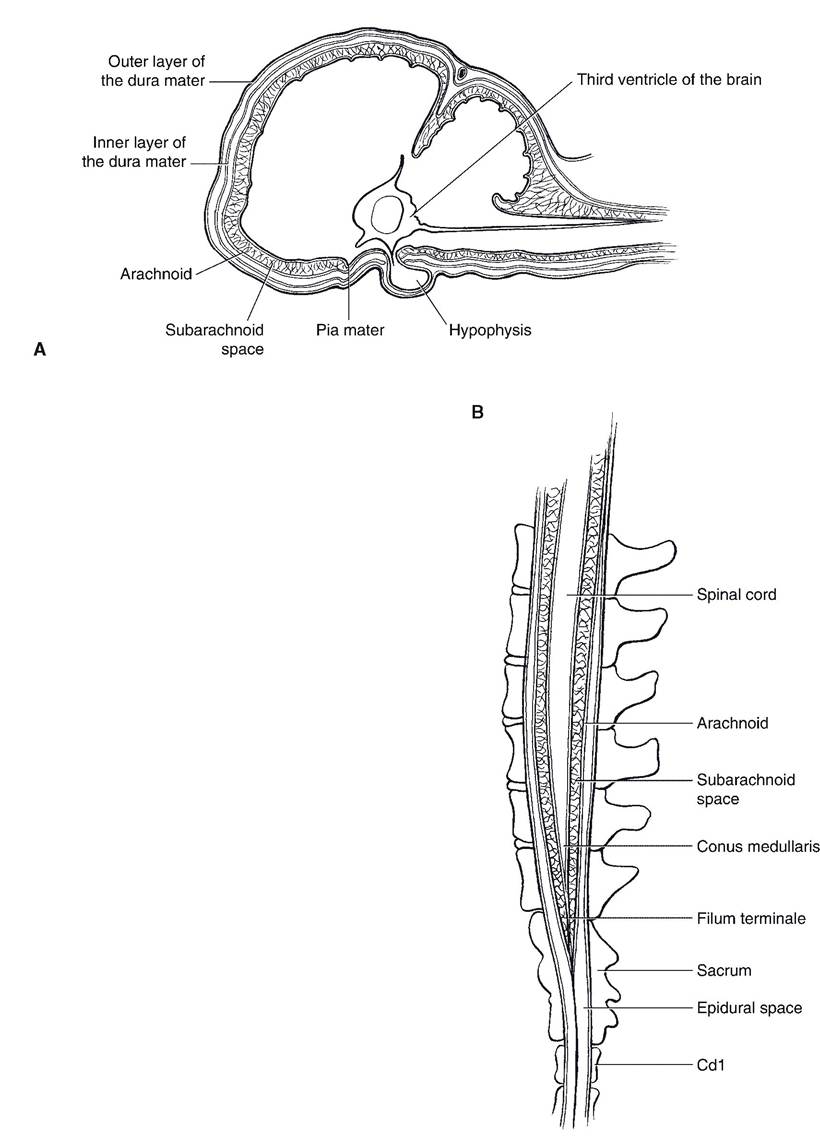
Figure 9-11. Meninges. A) Cranial meninges. B) Spinal meninges, depicted at the caudal end of the spinal cord. (Reprinted with permission of Wiley-Blackwell from Smith B.J. Canine Anatomy. Philadelphia: Lippincott Williams & Wilkins, 1999.)
and arachnoid constitute the leptomeninges (from the Latin word Iepto, delicate), reflecting their fine, delicate nature. The space between the two layers, bridged by arachnoid trabeculae, is the subarachnoid space. It is filled with CSF. It is from this space that CSF is collected when a spinal tap is performed.
The dura mater is the tough fibrous outer covering of the CNS. Within the cranial vault the dura mater is intimately attached to the inside of the cranial bones and so fulfills the role of periosteum. It also forms the falx cerebri, a median sickle-shaped fold that lies in the longitudinal fissure and partially separates the cerebral hemispheres. Another fold of dura mater, the tentorium cerebelli, runs transversely between the cerebellum and the cerebrum. In some locations within the skull, the dura mater splits into two layers divided by channels filled with blood. These dural sinuses receive blood from the veins of the brain and empty into the jugular veins. They are also the site of reabsorption of CSF through the arachnoid granulations back into the circulation.
The pia mater and the arachnoid of the spinal meninges are much as they are within the cranial vault. The dura mater of the spinal meninges, however, is separated from the periosteum of the vertebral canal by a fat-filled space, the epidural space.
It is into the epidural space that physicians and veterinarians introduce local anesthetics to produce anesthesia in the caudal parts of the body. This procedure, epidural anesthesia, is often done for obstetric procedures. One common application is the injection of anesthetic between the first and second caudal vertebrae of the ox as an aid to repairing and reducing uterine, vaginal, or rectal prolapses.
Spinal Cord
The spinal cord (Figs. 9-12 and 9-13) is the caudal continuation of the medulla oblongata. Unlike that of the cerebrum, the spinal cord’s gray matter is found at the center of the cord, forming a butterfly shape on cross-section. Fiber tracts, the white matter, surround this core of gray matter. A spinal cord segment is defined by the presence of a pair of spinal nerves. Spinal nerves are formed by the conjoining of dorsal and ventral roots, which come together as the nerve at the point where the axons exit and enter the vertebral canal.
Sensory neuronal cell bodies are present in aggregates, called dorsal root ganglia, lateral to the spinal cord. The neurons within these ganglia are pseudounipolar, and they give rise to processes that enter the dorsal horn of the
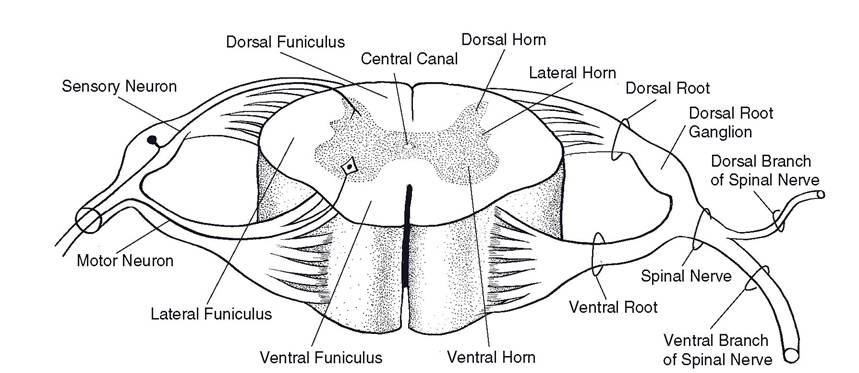
Figure 9-12. Cross-sectional anatomy of the spinal cord and spinal nerves.
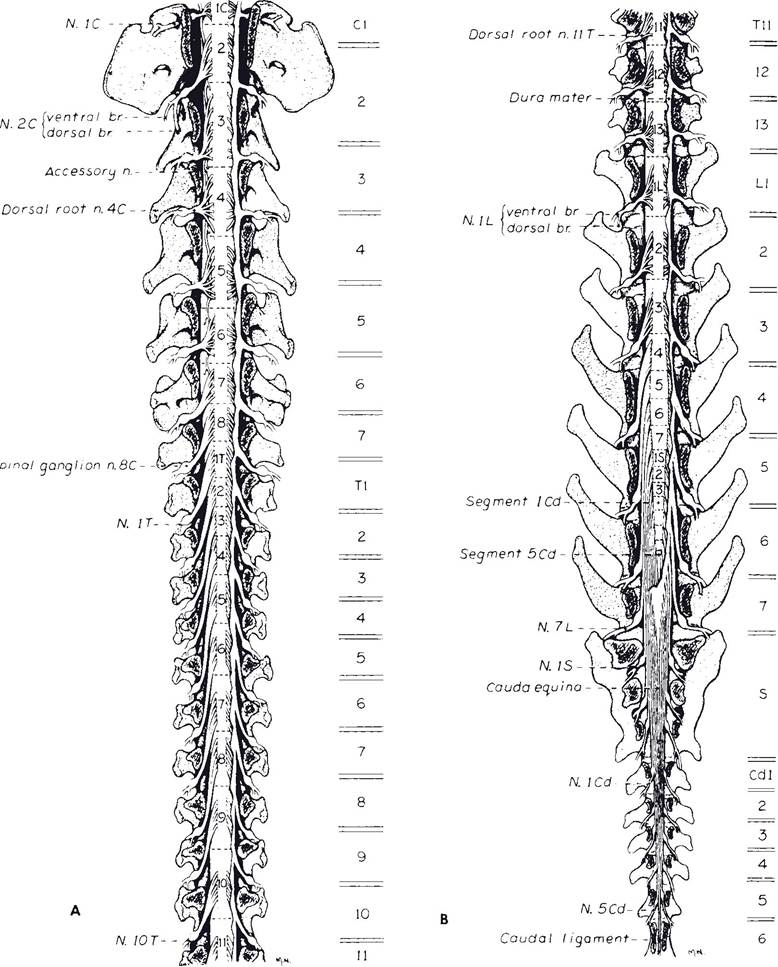
Figure 9-13. Spinal cord of the dog. View is from dorsal side, with the vertebral arches removed to expose the cord within the vertebral canal. Note the relation of the spinal nerves to the vertebral bodies. The spinal cord is shorter than the vertebral column, so that the more caudal spinal cord segments lie within vertebrae whose name and number are more cranial (e.g., spinal cord segment L6 lies within vertebra L4). The nerve roots are progressively longer in these more caudal parts of the vertebral column as they extend from the cord to the foramen of exit. (Reprinted with permission from Fletcher T.F. and Kitchell R.L. Anatomical studies on the spinal cord segments of the dog. AJ.VR. 1966;27:1762.)
spinal cord and others that unite with motor fibers from the ventral horn neurons to become the spinal nerve extending into the periphery. The processes that extend from the spinal nerve to the spinal cord constitute the dorsal root.
The ventral root of the spinal nerve consists of motor fibers that arise from the nerve cells primarily in the ventral horn of the spinal cord. The dorsal and ventral roots unite to form the spinal nerve close to the intervertebral foramen between adjacent vertebrae. The dorsal root ganglion is usually very close to this conjoining of dorsal and ventral roots; it frequently can be found just within the intervertebral foramen. In this location it is susceptible to compression when an intervertebral disk protrudes; to a large degree, it is this compression that causes the intense, electric pain associated with disk disease.
Tracts of the Spinal Cord. A tract is a bundle of functionally related axons in the CNS. Tracts that carry sensory information are ascending tracts, whereas those carrying motor commands are descending tracts. The white matter of the spinal cord in which the tracts are found can be roughly divided into three columns on each half of the cord: a dorsal funiculus (often called the dorsal column), a lateral funiculus, and a ventral funiculus.
Sensory Tracts. The dorsal funiculi contain afferent tracts that carry information about body position from joints, tendons, and muscles. This type of sense is called proprioception. Injury to this pathway produces uncoordinated, inaccurate movements, as the cortex lacks some of the information it needs to make ongoing adjustments in the planning and execution of voluntary movements. Such incoordination is sensory ataxia.
Proprioceptive information also ascends the spinal cord in several spinocerebellar tracts, located superficially on the lateral funiculus. As the name suggests, these tracts are headed for the cerebellum, where the proprioceptive information is used to help shape voluntary movements so that they are accurate and smooth.
Information about pain is carried by a large number of described pathways, but these are often grouped under the names spinothalamic tract or anterolateral system. This large group of fibers is found in a wide band through the lateral and ventral funiculi. Some of these tracts terminate in the brainstem, where they mediate reflexes associated with painful stimuli. Others make connections that alert the entire cortex and initiate aversive behaviors. Still others are relayed directly to the parts of the cortex that create conscious awareness of the painful stimulus.
Like pain, touch and temperature sense information is carried in a variety of ascending tracts. Some of these are found in the dorsal columns and others in the anterolateral system.
Motor Tracts. Motor systems are often functionally grouped into two main categories: a ventromedial motor system, largely located in the ventral funiculus, and a dorsolateral motor system, found in the dorsal part of the lateral funiculus.
The ventromedial motor system primarily is responsible for activity in the axial and proximal limb muscles, especially extensors and antigravity muscles. Activity in the tracts of this system assists with the support phase of gait, when limbs are in weight-bearing position with joints extended. A particularly noteworthy tract of the ventromedial motor system is the lateral vestibulospinal tract, which originates in the region of the pons and medulla.
The dorsolateral motor system is in many ways complementary to the ventromedial system. Dorsolateral tracts tend to control the muscles of the distal limb, especially the flexors. Activity here is important in the flexion or swing phase of gait, when limbs are lifted from the ground and advanced while flexed. In quadrupeds, the most prominent tract in the dorsolateral motor system is the rubrospinal tract, which arises from the midbrain. In primates, and most especially humans, the corticospinal tracts arising from the motor cortex of the cerebrum are especially well developed.