Cerebellar Histology and Phylogeny Give Clues to Cerebellar Function
The cortex throughout the cerebellum consists of three layers and only five types of neurons: stellate, basket, Golgi, granule, and Purkinje cells (Figure 12-2). The outermost layer is the molecular layer and consists primarily of granule cell axons, known as parallel fibers; dendrites of neurons located in deeper layers; and scattered inhibitory interneurons: the stellate and basket cells (Figure 12-3).
The middle Purkinje cell layer consists of the large cell bodies of Purkinje neurons, which have a flat but extremely expansive dendritic field oriented at right angles to the parallel fibers (see Figure 12-2). This arrangement, and because a parallel fiber contacts the dendrites of many Purkinje cells, is concordant with the cerebellums role in the timing of movement components. Axons of the Purkinje neurons go to the deep cerebellar nuclei, located outside of the cerebellar cortex. The Purkinje cells are the only output neurons of the cerebellar cortex and are all inhibitory, using gamma-aminobutyric acid (GABA) as their transmitter. They can inhibit the spontaneously active neurons of the deep cerebellar nuclei, whose axons leave the cerebellum. The innermost granule cell layer of the cerebellar cortex contains a vast number of small granule cells and occasional Golgi cells.The two primary groups of input axons to the cerebellum are the mossy fiber and climbing fiber axons (see Figure 12-2). Both are excitatory; they cause excitatory postsynaptic potentials (EPSPs) within the cerebellar cortex and, through collateral axons, within the deep cerebellar nuclei. The mossy and climbing fibers collectively carry information from components of the motor system hierarchy and from peripheral sensory receptors regarding the planning and execution of the movement. The shorter input/output circuit of the cerebellum consists of the climbing and mossy fiber stimulation to the deep cerebellar nuclei, whose output in turn leaves the cerebellum to modify components of the motor system hierarchy (Figure 12-4). However, the output of the deep cerebellar nuclei is itself modified by inhibition from Purkinje cell axons.
The Purkinje cell inhibition of deep cerebellar nuclei is based on the cerebellar cortex's own integration of mossy and climbing fiber inputs.Although the cortical Synaptology is understood, just how the cerebellum integrates movement feedback with the motor
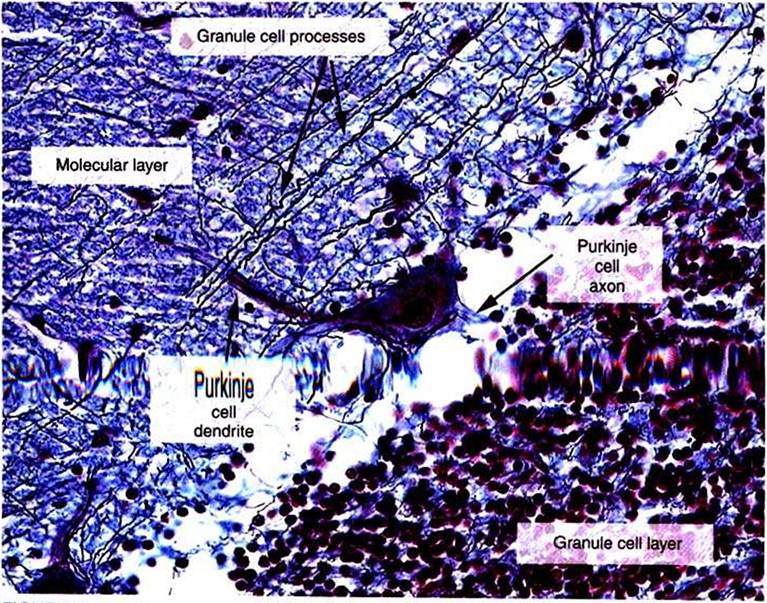
FIGURE 12-3 High-power photomicrograph of the three layers of the cerebellar cortex: granule cell layer, Purkinje cell layer (not labeled), and molecular IayenThe image provides a good example of how the parallel fibers, labeled "granule cell processes," cross the dendritic region of the Purkinje cell in the molecular layer. However, the elaborate branching pattern of the Purkinje cell dendrites is not apparent with this stain. (Image courtesy Dr.Tom Caceci, Department of Biomedical Sciences and Pathobiology, College OfVeterinary Medicine, VirginiaTech.)
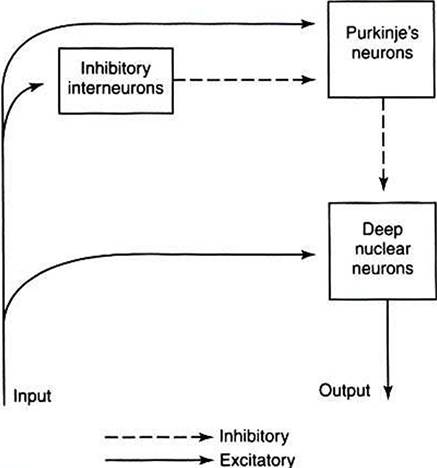
FIGURE 12-4 InputZoutput organization of the cerebellum.
plan and then modifies the output of the deep nuclear neurons is not clear. As noted earlier, because the histological appearance of the cortex is similar throughout the cerebellum, it seems likely that a similar underlying modulation process occurs regardless of the cerebellar region. However, regional variation of inputs from and outputs to different parts of the nervous system would render different motor results from different cerebellar regions. Therefore, it is useful to examine the three major, phylogenetically different divisions of cerebellum.
The cerebellum can be divided into three distinct regions from both a functional perspective and a phylogenetic perspective: the vestibulocerebellum, the Spinocerebelluni, and the Cerebrocerebellum (Figures 12-5 and 12-6).