Circulatory System
The circulatory system consists of a network of continuous tubes connected to a muscular pump. This system, as its name implies, is responsible for the transportation, distribution, and circulation of various chemicals throughout the body.
Examples of these activities include distribution of nutrients and transport of gases, hormones, and metabolites.Although the circulatory system is continuous, it can be examined as individual components, (1) the heart, (2) the arteries, veins, arterioles, and venules that are generally connected by capillary beds, and (3) the blood. Components of the blood are the fluid plasma and circulating elements, erythrocytes, leukocytes, and platelets. These elements play an important role in gas transport, immune reactions, and blood clotting.
A major function of capillaries is the diffusion of water and any dissolved substances between the circulatory system and tissues in which they occur. Under normal physiological conditions, blood elements, and proteins occurring in the blood are too large to pass across the capillary wall. The overall result is that a small amount of fluid remains in the surrounding tissues. This fluid is returned to the circulatory system by way of lymphatic vessels that enter the systemic circulation in the neck and shoulder region.
Although mammalian hearts are all very similar in morphology, in the discussion that follows, the anatomy described is that of the sheep. Our recommendation is that the heart be cut with a sharp kitchen knife into dorso- ventral halves preferably by your instructor.
THE HEART
External Anatomy
The mammalian heart consists of two muscular pumps, the right atrium and right ventricle, that receive and pump oxygen-poor blood from the body to the lungs and the left atrium and left ventricle that receive oxygen-rich blood from the lungs and pump it throughout the body [Figure 8-1 and Figure 8-2].
The heart is shaped somewhat like a blunt wedge, with its flattened base and more acute apex [Figure 8-1]. In the living mammal, including humans, the heart tilts to the left somewhat and projects into a membranous sac, the pericardium, in a potential thoracic space called the mediastinum. Therefore the left side of the heart lies against the diaphragm. The outer layer of the pericardium is fibrous and may be covered by the parietal pleura, as in the cat. Lining the pericardial cavity into which the heart projects is the serous membrane known as the parietal pericardium. The parietal pericardium reflects over the heart surface as the visceral pericardium.
The walls of the heart consist of three layers. The outer covering is a single layer of squamous epithelium that is really nothing more than the visceral layer of pericardium, also known as the epicardium or outermost layer of heart tissue. The muscular middle layer or myocardium makes up almost the entire tissue of the heart. The inner lining, continuous with the inner lining of circulatory vessels, also consists of a single layer of squamous epithelium known as the endothelium.
As you view the heart externally, perhaps your first impression is that the outer surface seems to be covered with an unusual amount of fat. Strange as it might seem, this is normal for the hearts of mammals.
The first determination that you should make is which side is dorsal and which is ventral. Remember that the heart as it rests in the normal living sheep's body has a dorsal and ventral aspect. In mammals, no matter how they walk, the pulmonary trunk, a major blood vessel that exits from the right ventricle, stretches from right to left across the ventral side of the heart [Figure 8-1]. Another vessel, the aorta, generally of greater diameter and slightly dorsal to the pulmonary trunk, also curves left. From the aorta branches the brachiocephalic artery. Carefully pick away fat between the pulmonary trunk and aorta until you discover a short ligamentous band connecting the two vessels.
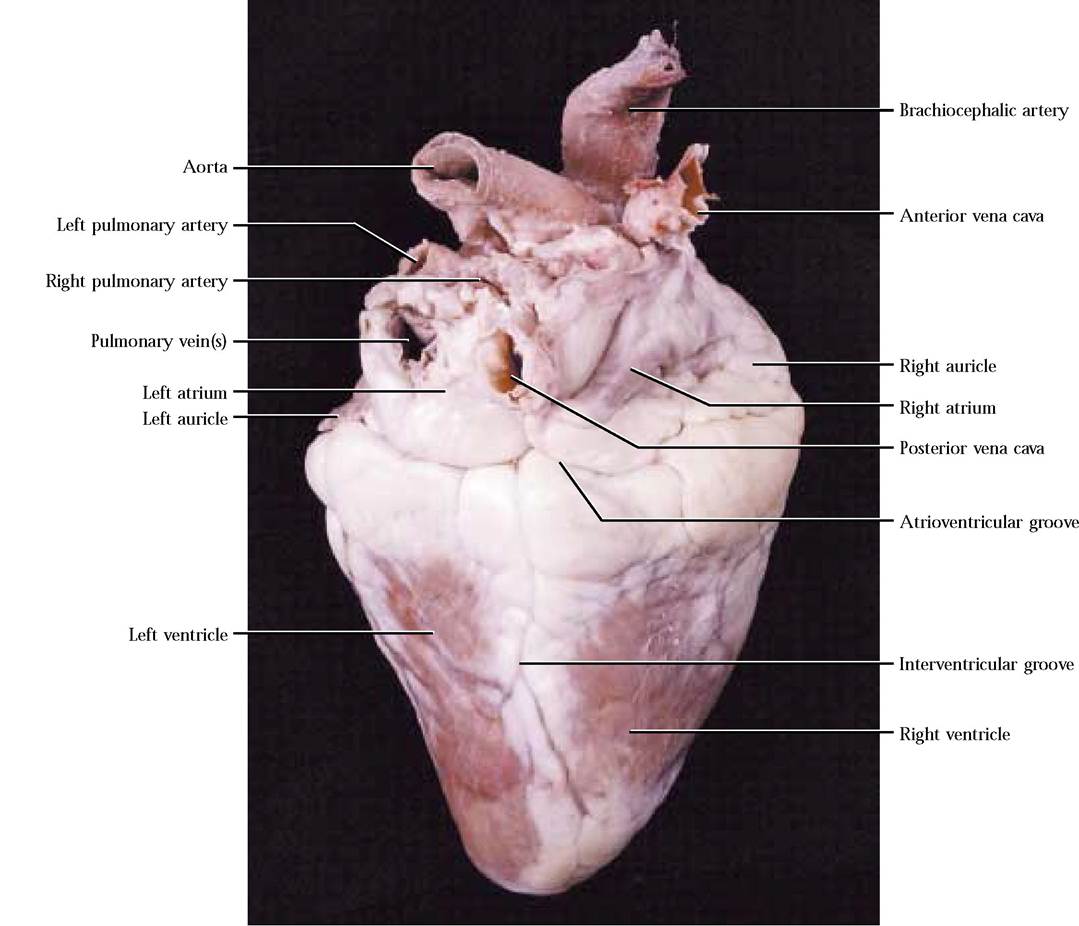
This represents the remnant of a fetal circulatory bypass, the ligamentum arteriosum, which allows most of the blood pumped into the pulmonary trunk during intrauterine life to be shunted into the systemic circulation and away from the nonfunctional lungs [Figure 8-1]. If you turn the heart over, you will probably notice that there are a greater number of vessels, the four pulmonary veins and the anterior and posterior vena cavas, associated with the dorsal side [Figure 8-2]. In some specimens, the dorsal vessels are sometimes cut very close to the surface of the heart, leaving only openings or holes.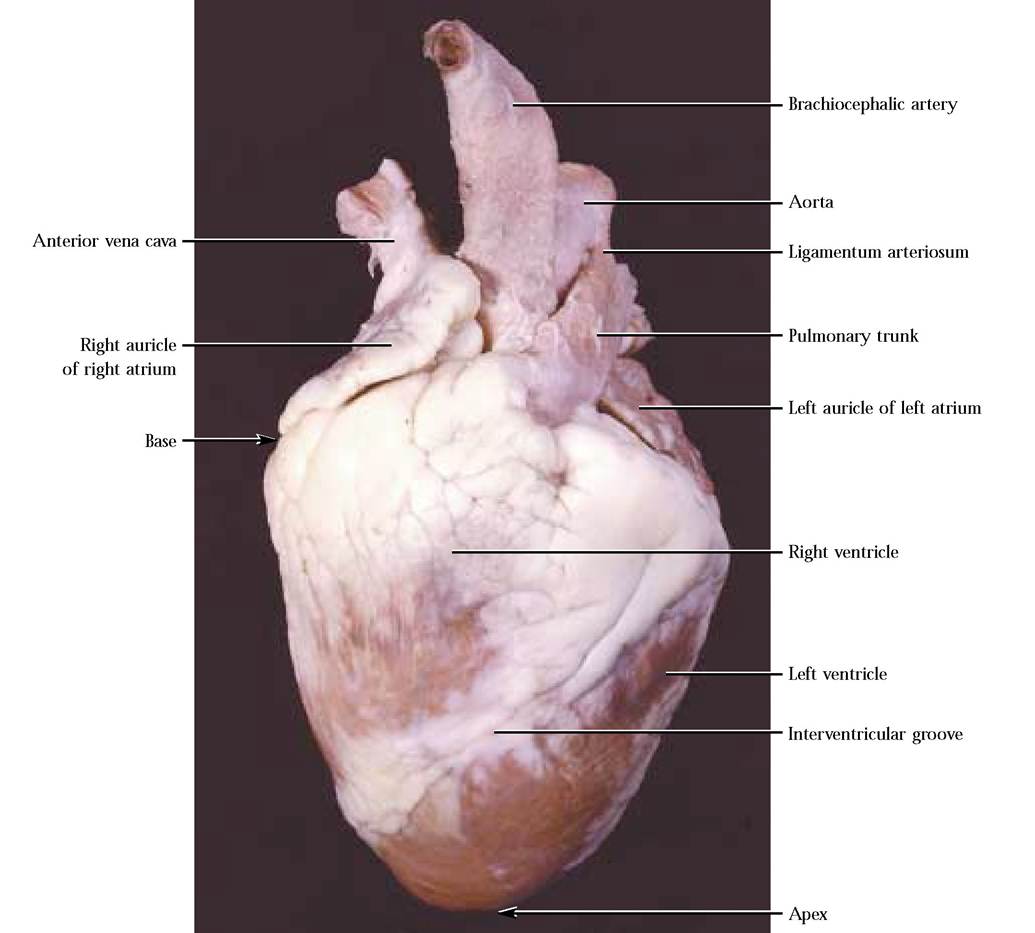
A shallow atrioventricular groove, or coronary sulcus, marks the separation of the heart into the two anterior atria and the two posterior ventricles [Figure 8-2]. Encircling the heart in this groove is the main venous drainage of the heart, the coronary sinus. Venous blood returns to the heart from heart muscle through this channel. Interventricular grooves on the ventral and dorsal surfaces demarcate the division of the ventricle into right and left chambers [Figure 8-1 and Figure 8-2]. Major coronary blood vessels lie in these grooves, generally buried in a considerable amount of fat. Note that these grooves are oriented almost parallel to the left margin of the heart, giving the appearance that the left ventricle is larger. This is not an optical illusion, but due to the greater thickness of muscle in the left ventricle to develop the pressures needed to pump oxygen-rich blood throughout the systemic circulation. This disparity in ventricular thickness is very obvious upon examination of the cut surface of the muscular walls of both chambers. Also obvious are two ear-like flaps, the auricles, attached to the atria [Figure 8-1 and Figure 8-2]. They increase the volume of each atrium somewhat.
Internal Anatomy
In our treatment of the internal morphology we will describe the anatomy with respect to the incoming blood flow through the right pump, followed by the outgoing blood flow through
the left pump.
Be aware that both pumps function at the same time.Blood enters the right atrium from the cranial region of the body by way of the anterior vena cava [Figure 8-3] and from the caudal region of the body via the posterior vena cava [Figure 8-3]. Locate the openings of these two major vessels in the inner wall of the right atrium. Notice that the atrial wall is thin compared to the ventricle. If there is a substantial section of these vessels remaining attached to your specimen, notice that the posterior vena cava curves along the dorsal aspect of the lower left side of the heart, turning cranially to enter the right atrium. Additionally, locate the opening of the coronary sinus [Figure 8-3] situated caudal to the opening of the posterior vena cava. Venous blood from the heart, itself, drains into the right atrium through this opening. The inner wall of the auricle appears honeycombed because of the presence of muscular ridges, the musculi pectinati [Figure 8-4]. These muscular ridges also extend into the atrium. A depression, somewhat lighter in color than the surrounding tissue, sits just ventral to the entrance of the posterior vena cava in the interatrial septum. This area, the fossa ovalis, represents the position of an opening between the right and left atria that functions as a shunt during embryonic development [Figure 8-3].
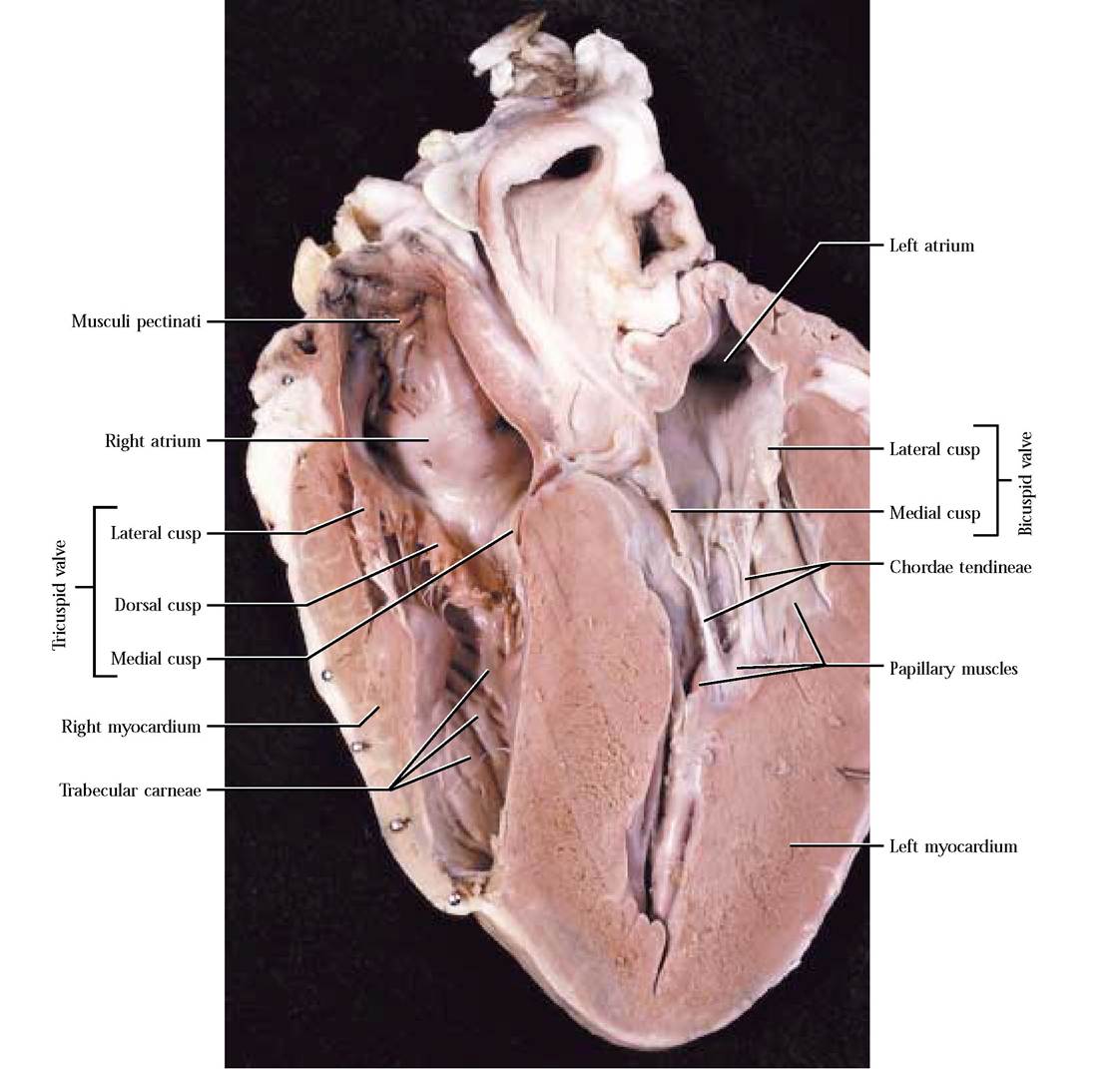
Blood flows from the right atrium into the right ventricle through the right atrioventricular or tricuspid valve, taking its name from three flap-like structures or cusps—the dorsal cusp attached to the dorsal wall, the lateral cusp attached to the lateral wall, and the medial cusp attached to the medial wall [Figure 8-3 and Figure 8-4]. Attached to the apex of each cusp are cord-like structures, the chordae tendineae, extending to cone shaped papillary muscles in the wall of the right ventricle [Figure 8-3 and Figure 8-4]. Because the tricuspid valve is open, most of the ventricular filling is passive, but the final ventricular volume results from atrial contraction.
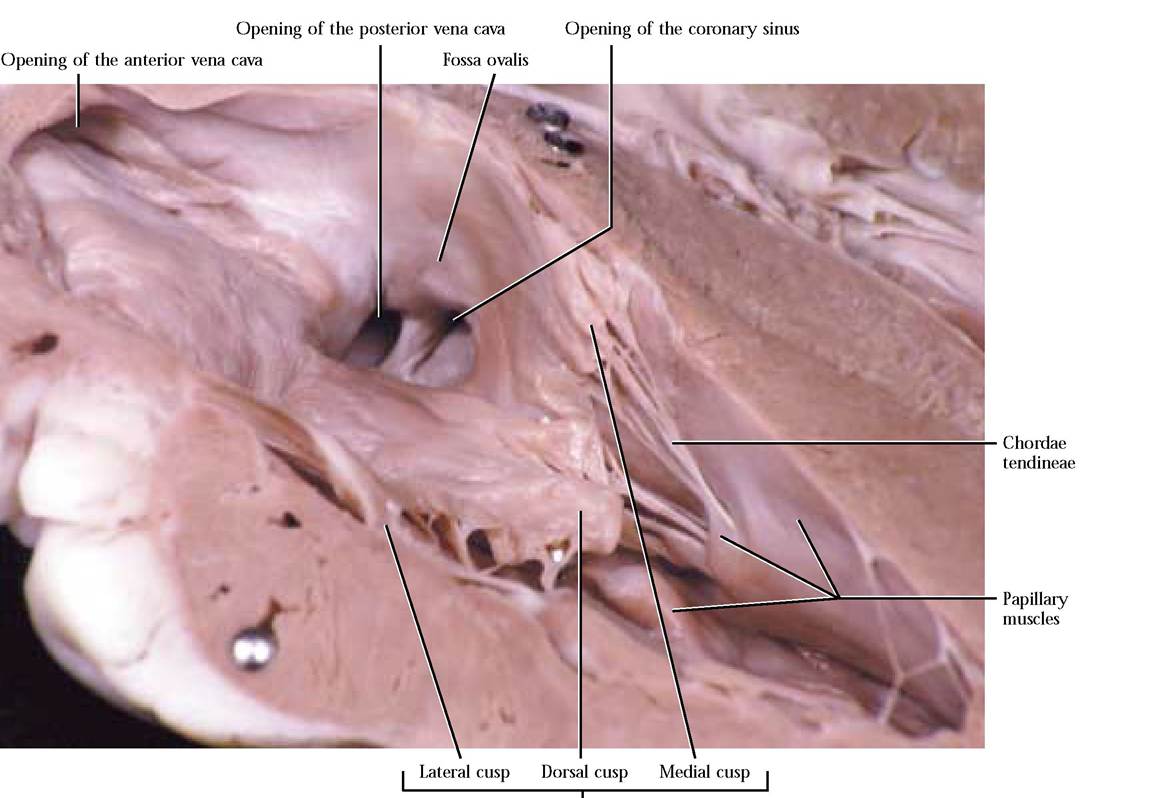
Tricuspid valve
As the right atrium relaxes and the right ventricle contracts, high pressure in the ventricle has a tendency to cause backflow of blood into the atrium causing the valve to slam shut, but the contraction of the papillary muscles pulling on the chordae tendineae prevent the eversion of the cusps into the atrium.
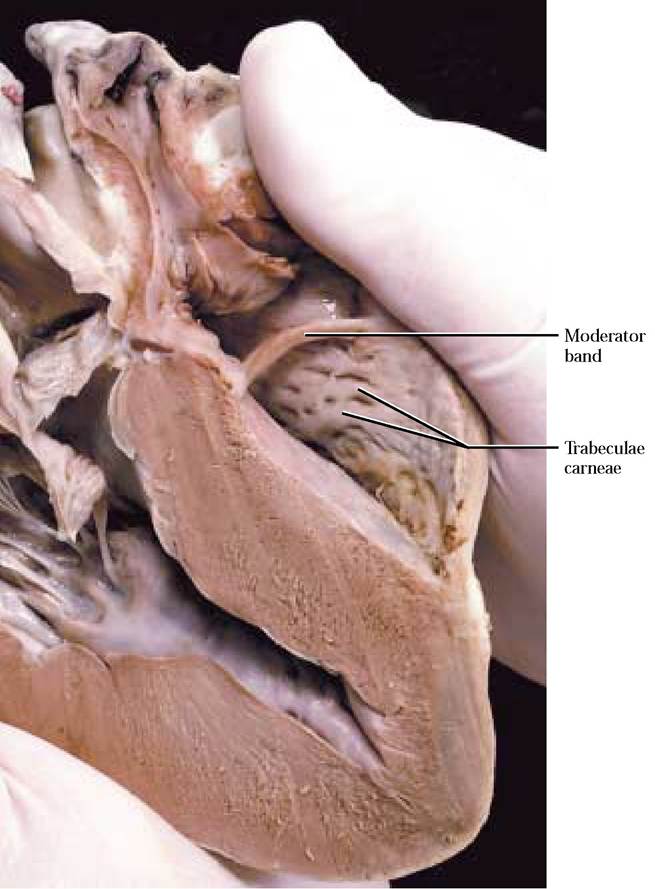
Notice that the inner walls of the ventricle are crisscrossed with muscular bands, the trabeculae carneae [Figure 8-4 and Figure 8—5]. A slender band of tissue, the moderator band, extends between the lateral and medial wall of the right ventricle [Figure 8—5].Note that the myocardium of the right ventricle is thinner than that of the left ventricle, since pressures are lower in the right ventricle because blood pumped by the right side travels only to the lungs located close to the heart [Figure 8-6].
From the right ventricle, blood is pumped through the pulmonary semilunar valve into the pulmonary trunk. Three membranous pockets, each in the shape of a half moon, form this valve. The decrease in pressure in the ventricle as it relaxes, causes the blood, under high pressure in the pulmonary trunk, to backwash into the ventricle. This backwash is prevented by blood filling the pockets of the semilunar valve which now slam shut. Blood is transported to the lungs where gas exchange occurs.
From the lungs, blood returns through the pulmonary veins to the left atrium. The two veins from each lung become confluent as they enter the atrium, sometimes giving the appearance of only two openings. Locate the openings of the pulmonary veins on the posterior aspect of the atrium [Figure 8-2]. Often, the pulmonary veins are removed close to the heart and appear only as holes in the left atrium.
The inner appearance of the left atrium is quite similar to the right atrium, with the exception that the musculi pectinati seem to be less extensive. However, they are prominent in the left auricle. As in the right pump, blood flows passively from the left atrium into the left ventricle, in this case, through the two cusps of the left atrioventricular or bicuspid valve. A medial and a lateral cusp are attached to those respective walls [Figure 8-6].
In a manner similar to the construction of the tricuspid valve, chordae tendineae extend between the margins of the cusps to papillary muscles protruding from the muscular walls of the ventricle. The final filling of the ventricle is accomplished by contraction of the left atrium.As the atrium relaxes and the ventricle contracts, the pressure of the engorged ventricle supercedes that of the atrium and blood backwash is again prevented when the cusps of the bicuspid valve slam shut. Contraction of the papillary muscles resulting in a pull on the edges of the cusps prevents eversion of the cusps into the left atrium.
The obviously thicker myocardium of the left ventricle is one of its most distinguishing features. Although it cannot be observed readily, an interventricular septum is present between the two ventricles, indicated externally by the interventricular grooves. Trabeculae carneae are again a feature of the inner surface of the chamber. Contraction of the left ventricle forces blood past the aortic semilunar valve into the aorta [Figure 8-6]. The morphology of this semilunar valve is identical to the pulmonary semilunar valve. As the ventricle relaxes, the blood pressure in the aorta exceeds that of the ventricle and blood backwashes toward the ventricle, filling the membranous pockets causing them to slam shut.
Continuous pumping of the heart throughout the life of a mammal demands a constant
supply of highly oxygenated blood to this hard working muscular organ. This requirement is met by delivery of the most highly oxygenated blood by way of the left and right coronary arteries that originate immediately above the aortic semilunar valve. Examine the medial and lateral walls of the aorta where you will observe the openings of these two vessels [Figure 8-6]. You may observe a number of small openings that represent coronary blood vessels.
With the exception of the moderator band, the anatomy of the human heart is very similar to the sheep.
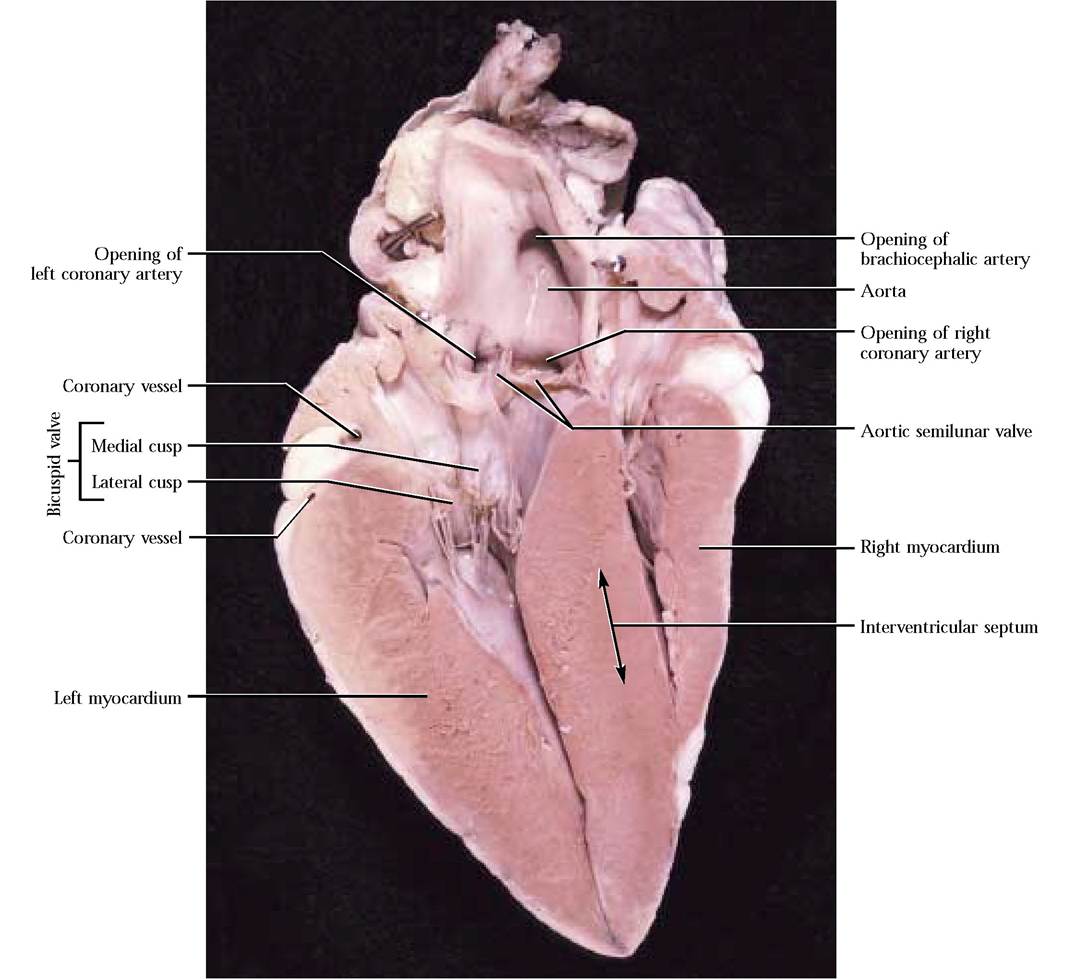
FIGURE 8-6 Left atrioventricular valve.
BLOOD VESSELS
The arteries, veins, arterioles, and venules that are generally connected by capillary beds are the circulatory channels through which blood moves. In this section, you will be dissecting the major blood vessels.
When you were in grade school, you learned about the primary colors, red, blue, and yellow. In a triply injected specimen, these same colors appear as injected latex in the vessels. Arteries are red, veins are blue, and the hepatic portal system consisting of the veins of most of the abdominal organs have been injected with yellow latex.
Pulmonary Circulation
This circuit includes arterial vessels carrying oxygen poor blood to the lungs and venous vessels carrying oxygen rich blood back to the heart. Take note that these vessels follow the anatomical rules where blood is carried back to the heart by veins and away from the heart by arteries. However, in contrast to the blood vessels of the rest of the body of an adult, the oxygen level of the blood is low in the pulmonary arteries and high in the pulmonary veins.
In order to observe these vessels, it is necessary to cut the pericardial sac with a pair of scissors. Identify the pulmonary trunk of the cat exiting from the right ventricle.
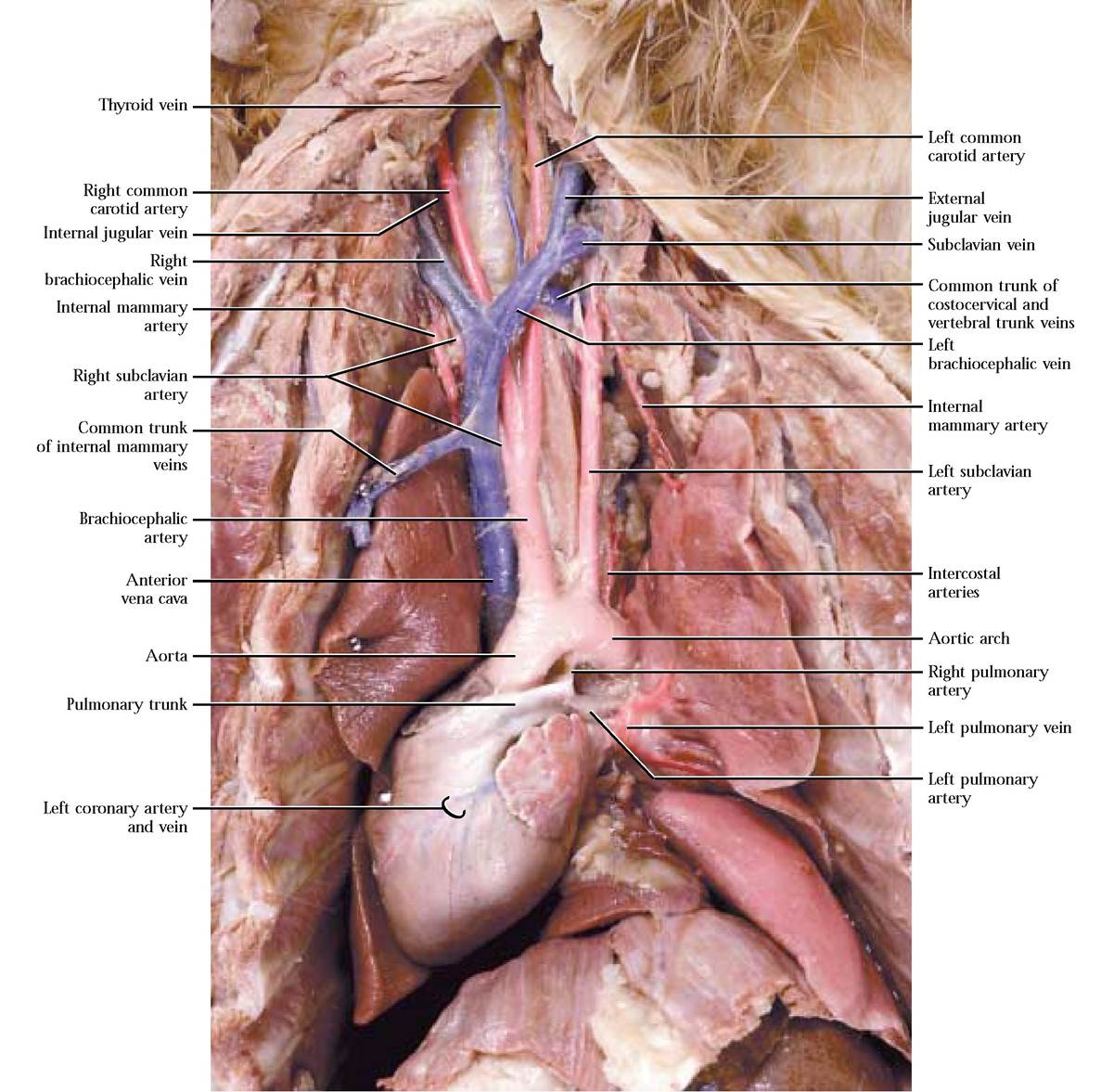
Dorsal to the heart the pulmonary trunk bifurcates into a right and left pulmonary artery [Figure 8—7]. Trace these vessels into the root of their respective lungs where they further branch, supplying the lobes of the lungs. In the root, identify the pulmonary veins as they leave the lungs and enter the left atrium [Figure 8-7]. Notice that the pulmonary veins contain red latex. The pulmonary circulation in humans is very similar.
Systemic Circulation
The architecture of the circulatory system exhibits a consistent pattern of divergence or branching in the arterial portion to supply the tissues of the body and convergence or confluence in the venous portion of the system to drain those tissues. Another common theme is that arteries and veins are paired when organs supplied or drained by them are paired, e.g., each kidney is supplied by an artery and a vein. You will also notice that a vessel that is continuous from its origin to its destination may carry different names as it passes from one area to another, e.g., the subclavian becomes the axillary as it enters the armpit but then is called the brachial as it enters the arm, and so on. This is quite common in the terminology of the circulatory system. In order to identify blood vessels correctly, it is necessary to trace them to or from the tissues that they supply.
The systemic circulation carries oxygen-rich blood from the heart to the body in arteries and returns oxygen-poor blood back to the heart in veins. To appreciate the enormity and complexity of the circulatory system, we will begin our exploration of the arterial component of this circuit by identifying the aorta as it leaves the left ventricle ventrally and curves dorsally to the left as the aortic arch [Figure 8—7]. The aorta then continues posteriorly, passing through the diaphragm into the peritoneal cavity, generally following the mid-dorsal line of the body. The portion of the aorta in the thoracic region may be identified as the thoracic aorta and that part in the peritoneal area may be identified as the abdominal aorta.
Blood Vessels of the Head, Neck, Chest and Forelimb
As the aorta begins to curve dorsally, identify the brachiocephalic artery followed by the left subclavian artery branching from the arch, supplying the neck and head and the forelimbs [Figure 8-7]. The brachiocephalic a., at about the level of the second rib, subdivides into the right subclavian artery, the right common carotid artery, and the left common carotid artery [Figure 8-7].
Four small arteries branch from each subclavian a. before it emerges from the body wall. In general, dissection of this portion of the circulatory system is minimal and consists mainly of picking connective tissue and fat from the surface of the vessels and nerves. As usual, this activity is to be done very carefully. Please take note that nerves often accompany the blood vessels and appear as shiny cream colored strands of tissue. These can be destroyed easily— be careful! The first or internal mammary artery leaves ventrally and enters the ventral thoracic body wall. As it courses posteriorly it gives off branches to nearby muscles, pericardium, mediastinum, and diaphragm and then anastomoses with the caudal epigastric arteries (described later). The next, the vertebral artery, arises as a dorsal artery, continues cranially and dorsally, enters the transverse foramen of the sixth vertebra, giving off branches to nearby neck muscles and spinal cord segments as it passes through the transverse foramina of the cervical vertebrae. As it nears the foramen magnum, it joins with the left vertebral artery to form the basilar artery that courses along the mid-ventral aspect of the medulla oblongata. The costocervical artery also emerges from the dorsal surface of the subclavian a., curves dorsally, and subdivides to send branches to neighboring neck, back, and intercostal muscles. The last of the subclavian a. branches, the thyrocervical artery leaves the anterior aspect of that vessel, travels anteriorly and laterally and as it approaches the shoulder becomes known as the transverse scapular artery. These vessels supply blood to adjacent muscles of the region, particularly the shoulder [Figure 8-8].
As the subclavian artery curves around the first rib and enters the axilla it becomes known as the axillary artery. From this artery, several others branch. The ventral thoracic artery emerges from the ventral surface of the axillary a. and courses posteriorly to supply pectoral muscles. Notice that the anterior ventral thoracic nerve of the brachial plexus accompanies this artery. A short distance laterally, the long thoracic artery branches and with the posterior ventral thoracic nerve of the brachial plexus supplies the pectoral muscles, thoracic mammary glands, and the latissimus dorsi muscle. The relatively large subscapular artery leaves the axillary as it nears the shoulder. The posterior humeral circumflex artery usually branches from the subscapular and accompanied by the axillary nerve of the brachial plexus, travels beneath the biceps brachii muscle carrying blood to several muscles of the medial aspect of the scapula, the shoulder, and triceps brachii muscle. A second branch of the subscapular a., the thoracodorsal artery, arises dorsal to the brachial plexus, giving off branches to the teres major and latissimus dorsi muscles [Figure 8-8]. The subscapular a. continues deep to supply dorsal shoulder muscles.
Beyond this point, the axillary a. becomes known as the brachial artery as it passes into the arm. The anterior humeral circumflex artery arises almost immediately as a branch of the brachial a. to bring blood to the biceps brachii muscle. At about the midpoint of the biceps brachii, the deep brachial artery originates and with the radial nerve of the brachial plexus passes under the belly of that muscle to supply blood to the triceps brachii, latissimus dorsi, and other muscles in the area. Occasionally, the deep brachial a. originates from the subscapular a. A number of muscular branches emerge from the brachial a. as it runs parallel with the median nerve of the brachial plexus until it nears the elbow where a pair of arteries, the radial collateral and ulnar collateral, are given off. The radial collateral artery turns laterally and supplies the arm extensor muscles while the ulnar collateral artery turns medially and supplies blood to the muscles of the elbow [Figure 8-8]. With the median
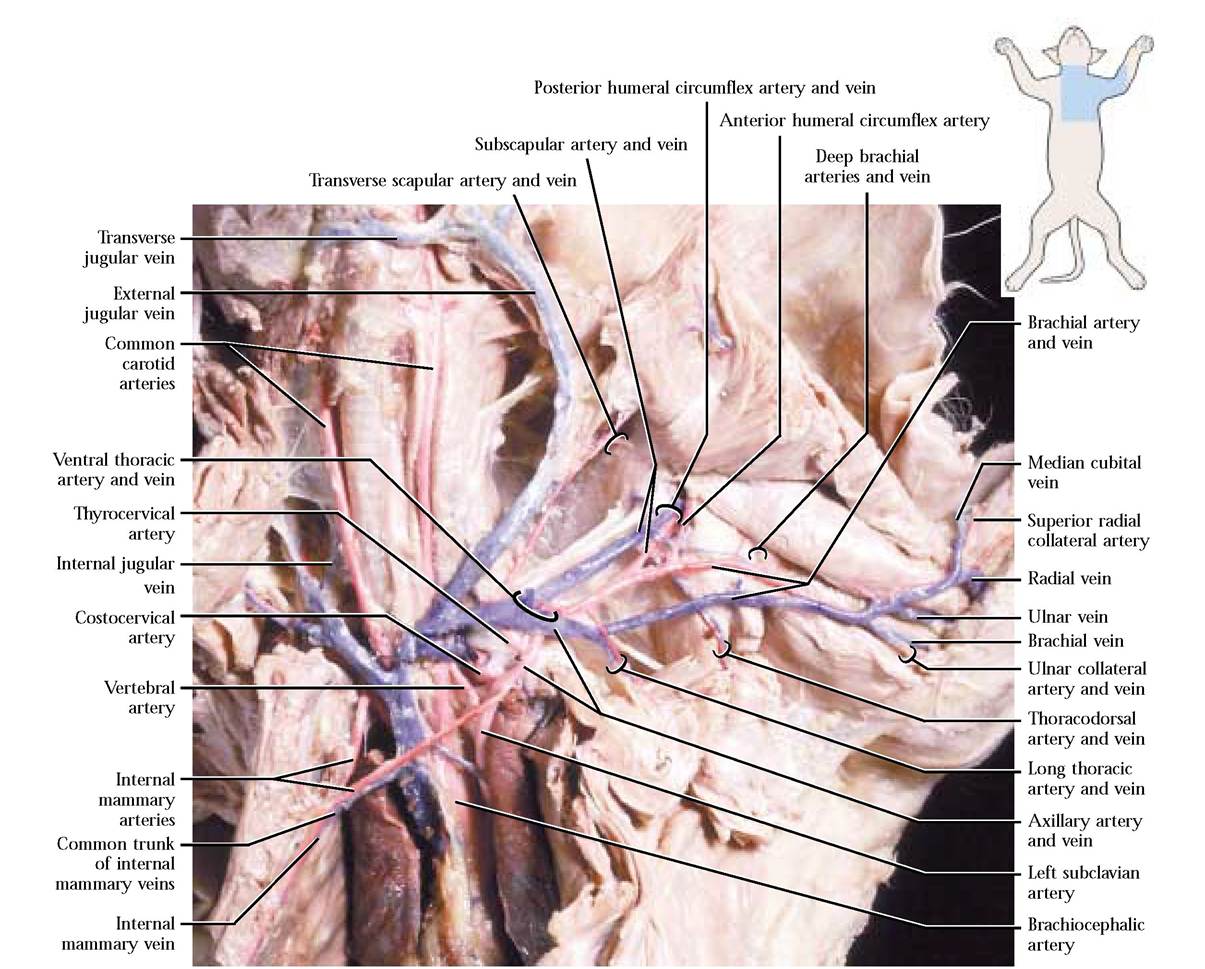
FIGURE 8-8 Vessels of the thoracic and brachial area.
nerve the brachial a. passes through the Supracondyloid foramen. Below this point it subdivides into an ulnar, a radial, and an interosseus artery.
The common carotid artery can be seen closely associated with the internal jugular vein which may not be well injected, the vagus nerve and sympathetic trunk and is bound to them by a fibrous sheath. Follow the common carotid cranially to the level of the thyroid gland and find a medial branch, the cranial thyroid artery, supplying the thyroid gland, parathyroid glands and neck muscles and directly opposite it, find the muscular branch passing laterally carrying blood to deep neck muscles. The next branch is the laryngeal artery that arises medially, supplying muscles in the laryngeal area. A variable number of branches may originate from the lateral side carrying blood to muscles and lymph nodes in the area. To view the cranial branches of the common carotid a., cut the sternomastoid and cleidomastoid muscles at the sternal end and reflect them. The hypoglossal nerve runs across the surface of the common carotid a. just anterior to the next pair of vessels that branch from it and serves as a landmark for the occipital and internal carotid arteries [Figure 8-9]. These vessels are generally very small, hairlike branches and may leave the dorsal part of the common carotid individually or may leave as a common stem that subsequently branches into two arteries. Great care must be exercised when probing for them because there is a great deal of connective tissue in the area and they are often not much more than 2-3 mm apart. The occipital a. originates first (more posteriorly) and passes caudal to the tympanic bulla carrying blood to the neck and occipital region. The internal carotid a. passes cranial to the tympanic bulla and transports blood to the base of the diencephalon, and along with several other arteries contributes to the major arterial blood supply of the brain in the form of the Circle of Willis.
The common carotid now continues as the external carotid artery following the branching of the internal carotid a. A large branch, the lingual artery, leaves the external carotid and carries blood to muscles of the hyoid and pharynx while continuing into the tongue as its major supply. The more dorsal external maxillary artery branches from the external carotid and carries blood to the outer facial region. As the external carotid nears the posterior boundary of the masseter muscle, the posterior auricular
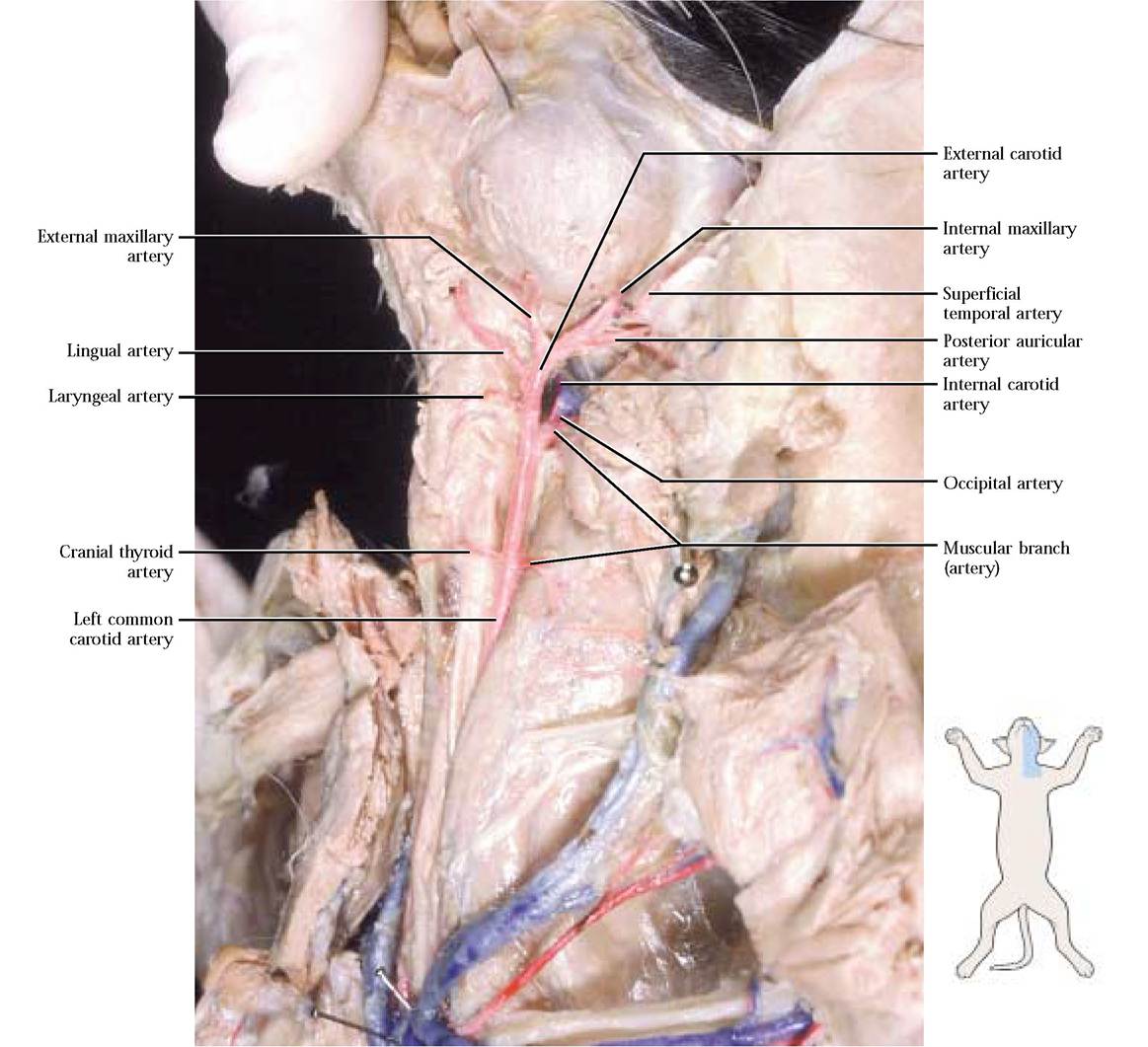
FIGURE 8-9 Branches of the left common carotid artery.
artery emerges and supplies the outer ear area [Figure 8-9]. The external carotid a. continues along the margin of the masseter, gives off the superficial temporal artery that delivers blood to the masseter and outer ear and continues deep to the masseter as the internal maxillary artery that supplies internal structures and tissues associated with the upper and lower jaw, nose, eyes, and surrounding regions [Figure 8-9]. In order to expose these vessels it will be necessary to carefully dissect some of the surrounding tissues. The branching pattern of the left and right sides is similar.
In the human, the branching pattern of the aortic arch is different. The first vessel, the brachiocephalic artery gives rise to the right common carotid and right subclavian arteries. The left common carotid and left subclavian arteries arise independently from the arch. The internal carotid artery is considerably more robust approaching the size of the external carotid in the human with several branches and serving as a major blood supply to the brain and regional organs. The arterial branches of the rest of this region are similar to the cat.
For the most part, arteries and veins lie in close proximity and as an artery is identified, the corresponding vein, often with the same name should be identified. In general, the venous system parallels the arterial system in this portion of the body with the following exceptions. Tributaries draining the brain, auricular, labial, lingual, palatal, and dental regions of the head form anterior and posterior facial veins that coalesce to form the external jugular vein. In the hyoid region, a transverse jugular vein draining that area connects the anterior facial veins. As the external jugular v. approaches the heart, it is joined by the transverse scapular vein draining the shoulder region. The brain and surrounding tissues are drained by tributaries of the internal jugular vein joining the external jugular [Figure 8-7, Figure 8-8, and Figure 8-10]. Notice that this vein lies close to the trachea and it may not be well injected.
Drainage of the forelimb involves deep and superficial vessels. The superficial veins include the cephalic vein that drains the lateral aspect of the forelimb and is connected to the brachial vein by the median cubital vein [Figure 8-11]. The cephalic v. continues superficially to join with the posterior humeral circumflex v. and the transverse scapular v. The deep veins of the forelimb parallel the arterial circulation.
The brachial-axillary-subclavian vein joins the external jugular v. to form the brachiocephalic vein. In contrast to the arterial supply, there are two brachiocephalic veins. The costocervical and vertebral veins merge to form a common trunk that empties into the brachiocephalic vein. Note that there is no thyrocervical vein. The left and right brachiocephalic veins join to form the anterior vena cava. The left and right mammary veins merge to form a common trunk and join the ventral surface of the anterior vena cava [Figure 8-7]. Although the pattern of the venous and arterial circulation differs somewhat, the regions served are similar.
Venous drainage of the head and neck region in the human is similar with these exceptions: the internal jugular vein is larger than the external jugular vein with many interconnecting channels between them. In contrast to the cat, the brachiocephalic veins are formed by the confluence of the internal jugular and the subclavian veins. The vertebral and external jugular veins join the subclavian independently.
Examine the inner thoracic wall and observe the segmental intercostal arteries and veins that are associated with the blood supply to the tissues between the ribs. The fairly large azygous vein runs along the right side of the aorta in the thoracic region and receives blood from most of the intercostal veins on both sides [Figure 8-12]. It empties into the dorsal region of the anterior vena cava. The origin of the azygous v. is actually in the dorsal abdominal area but it is best seen in the thoracic region and therefore is discussed here.
In humans, not only is there an azygous vein on the right side but also a hemizygous vein on the left side with a number of interconnecting vessels. An accessory hemizygous vein also exists.
Blood Vessels of the Abdominal Region
Among most vertebrates and in all mammals, as the aorta travels through the abdominopelvic region, it gives rise to an almost universal distribution of three unpaired major arterial branches, the coeliac artery, the cranial mesenteric artery and the caudal mesenteric artery, supplying the visceral organs [Figure 8-13]. To observe the complex subbranching of the coeliac a., carefully cut through the diaphragm to the esophagus, avoiding vessels associated with the diaphragm. Free the esophagus from connective tissue anchoring it in place and cut across the esophagus at a point about 2 cm anterior to the stomach. Pull the stomach ventro- medially. To begin the dissection of the coeliac and its branches, first locate the coelic a., then carefully pick away the connective tissue to expose its three major branches. The left gastric artery leaves the coeliac a. carrying blood to the lesser curvature of the stomach. Continue to pick away the connective tissue to completely expose the many smaller branches to the stomach. The hepatic artery branches from the coeliac a. and ascends toward the liver where it subdivides into several hepatic branches that supply the lobes of the liver and the cystic artery lying on the surface of the cystic duct. Use great care in exposing these delicate branches. As the hepatic artery travels toward the liver, observe a short gastroduodenal artery curving posteriorly and almost immediately giving off 1-3 delicate pyloric artery (ies) that supplies blood to the pyloric region of the stomach continuing along the lesser curvature of the stomach and anastomosing with the left gastric a. A few millimeters from the pyloric a.,
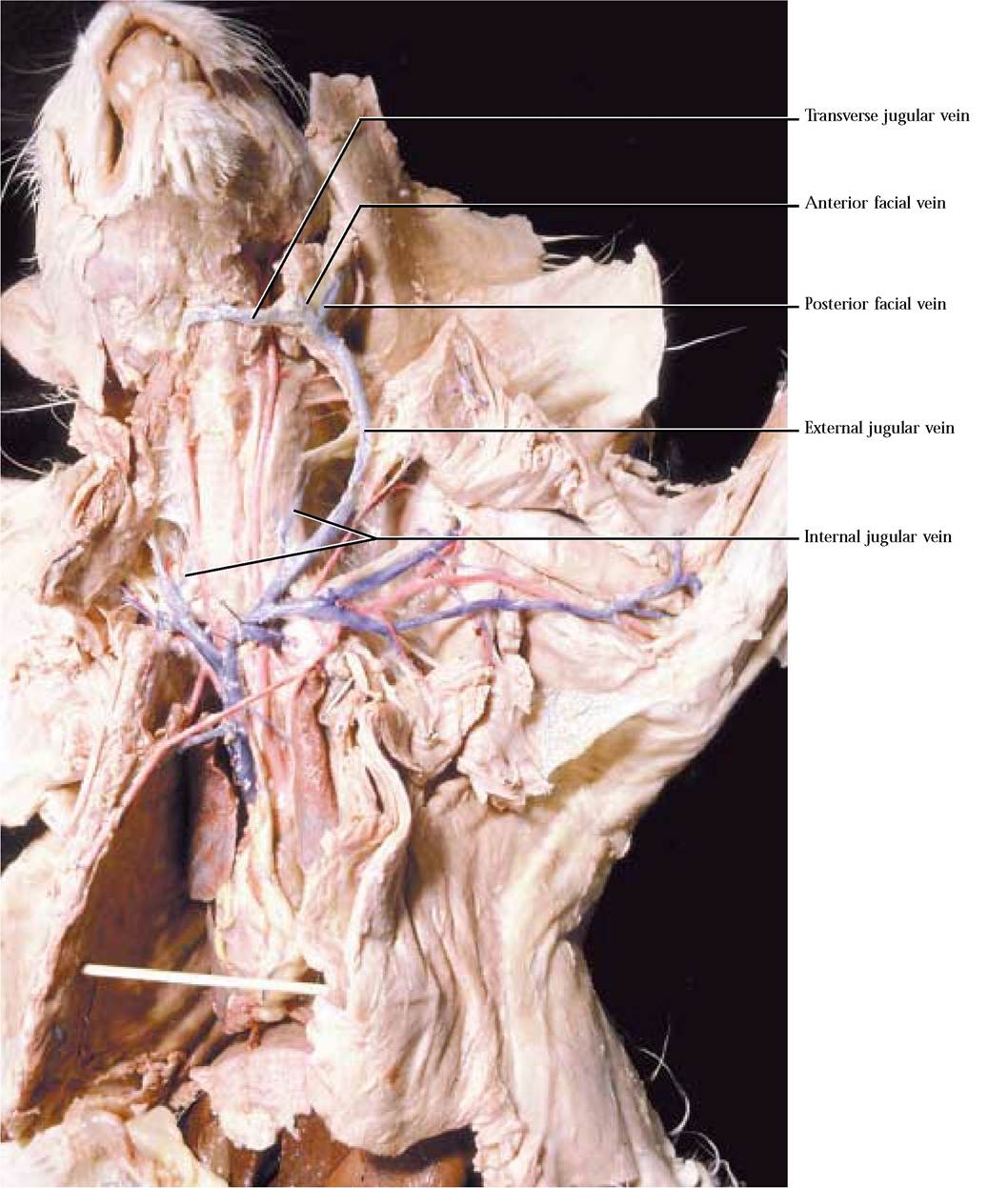
FIGURE 8-10 Veins of the head, neck, and thorax.
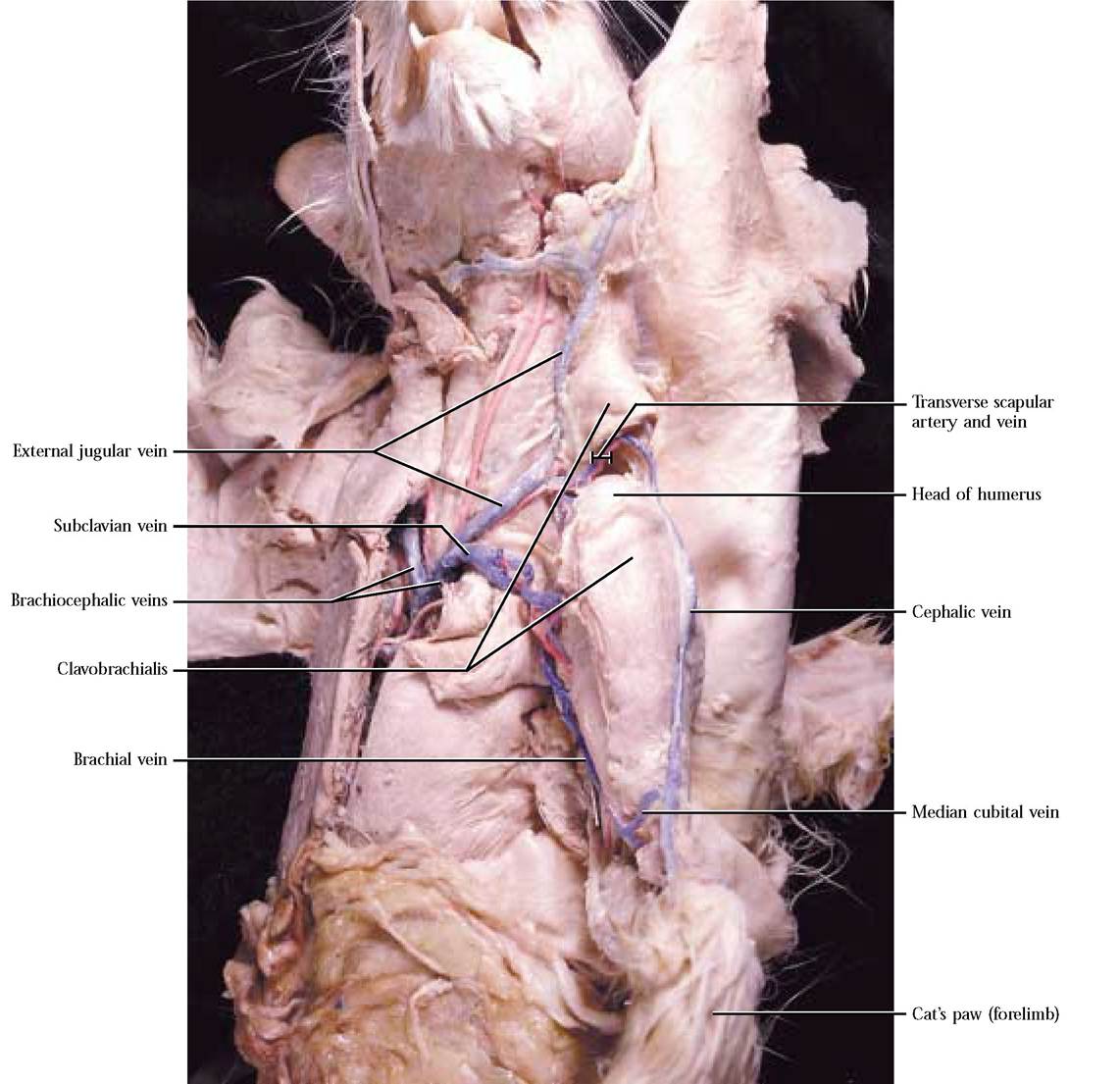
FIGURE 8-11 Superficial veins of the forelimb.
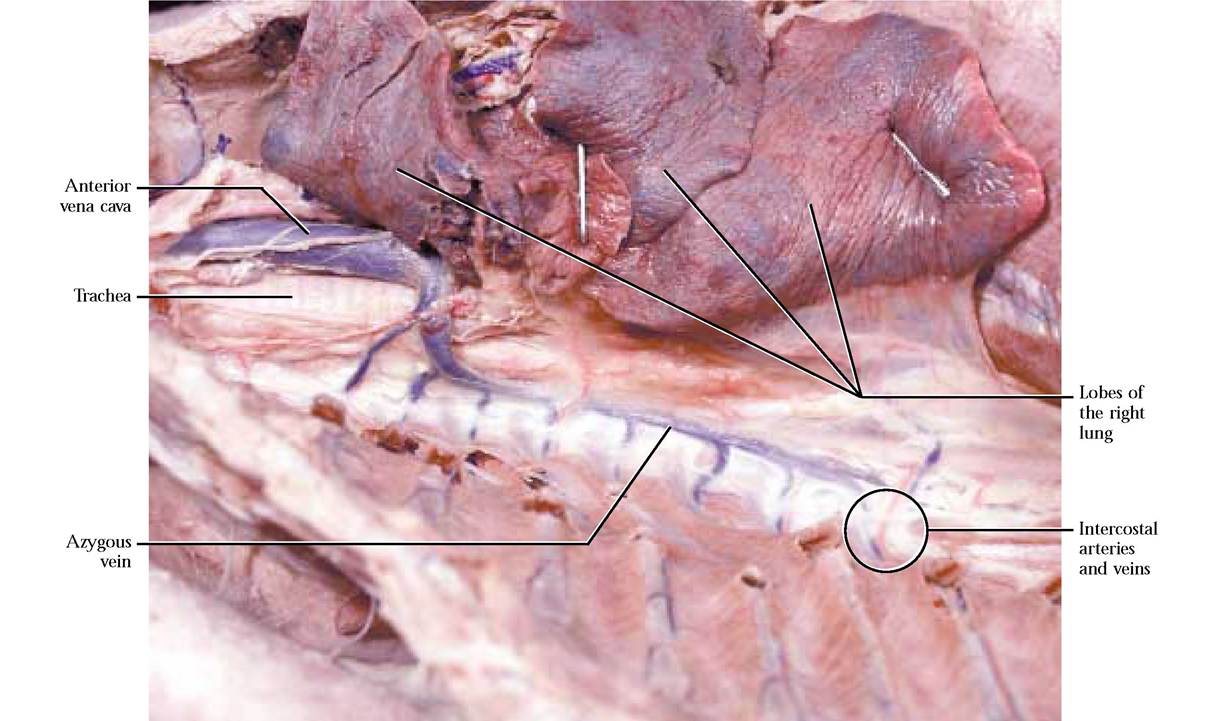
FIGURE 8-12 Azygous vein.
the gastroduodenal a. bifurcates into the right gastroepiploic artery supplying the greater curvature of the stomach and greater omentum and the anterior pancreaticoduodenal artery supplying the head of the pancreas and the duodenum [Figure 8-14B]. These arteries are generally more robust than the pyloric a., but you must still exercise great care in exposing them. The largest and most obvious branch of the coeliac, is the splenic artery leading toward the spleen. Notice the numerous small branches supplying the tail of the pancreas and the body of the stomach. The main portion of the artery travels along the length of the spleen giving off an anterior splenic artery and continuing as the posterior splenic artery [Figure 8-14A and Figure 8-14B]. A continuation of the posterior splenic a., the left gastroepiploic artery follows along the greater curvature of the stomach and anastomoses with the right gastroepiploic a. [Figure 8-16].
Although the description of the coeliac a. and its branches is applicable to most mammals, including the human, there may be variations in the pattern with additional small branches occurring.
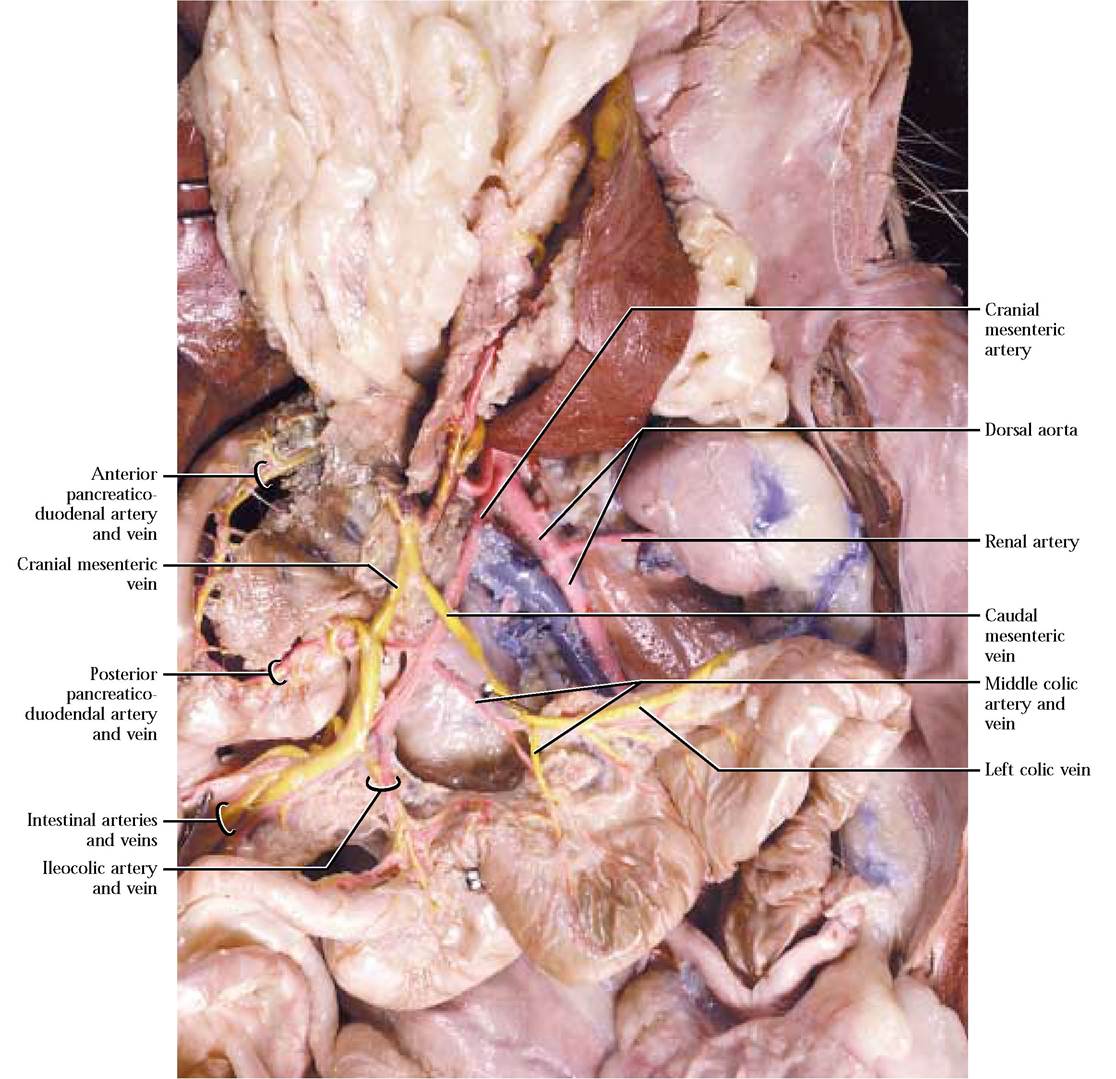
The second unpaired somewhat larger major abdominal artery is the cranial mesenteric artery, extending a greater distance than the coeliac a. before supplying blood to the intestines and pancreas. The first branch is the posterior pancreaticoduodenal artery supplying the pancreas and duodenum and anastomosing with the anterior pancreaticoduodenal a. [Figure 8-15]. Carefully expose this artery by picking away the connective and pancreatic tissue along the duodenal border. Follow the cranial mesenteric artery as it subdivides into numerous intestinal arteries, supplying the jejunum and ileum [Figure 8-15]. The next branch, the ileocolic artery, supplies the terminal portion of the ileum, the cecum, the ascending colon, and proximal portions of the transverse colon. Occasionally, a separate right colic artery, branching from the cranial mesenteric a., supplies the ascending and proximal portions of the transverse colon. The last branch of the cranial mesenteric a. is the middle colic artery supplying the transverse and descending colon [Figure 8-15]. Continuity among these colic vessels is the result of anastomoses. Again, the branching pattern of the cranial mesenteric artery of the cat and human is very similar.
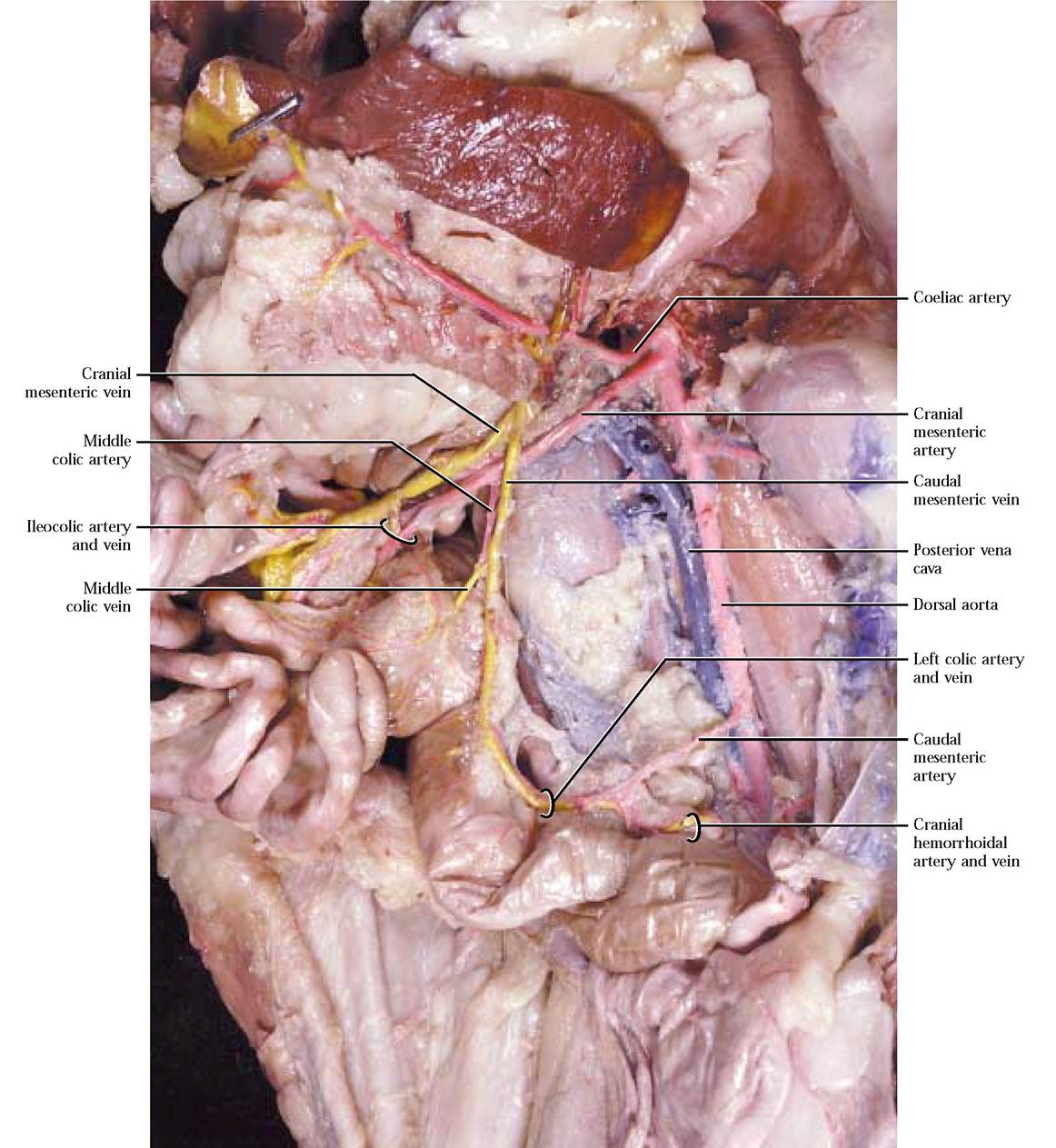
FIGURE 8-13 Distribution of the three abdominal arteries.
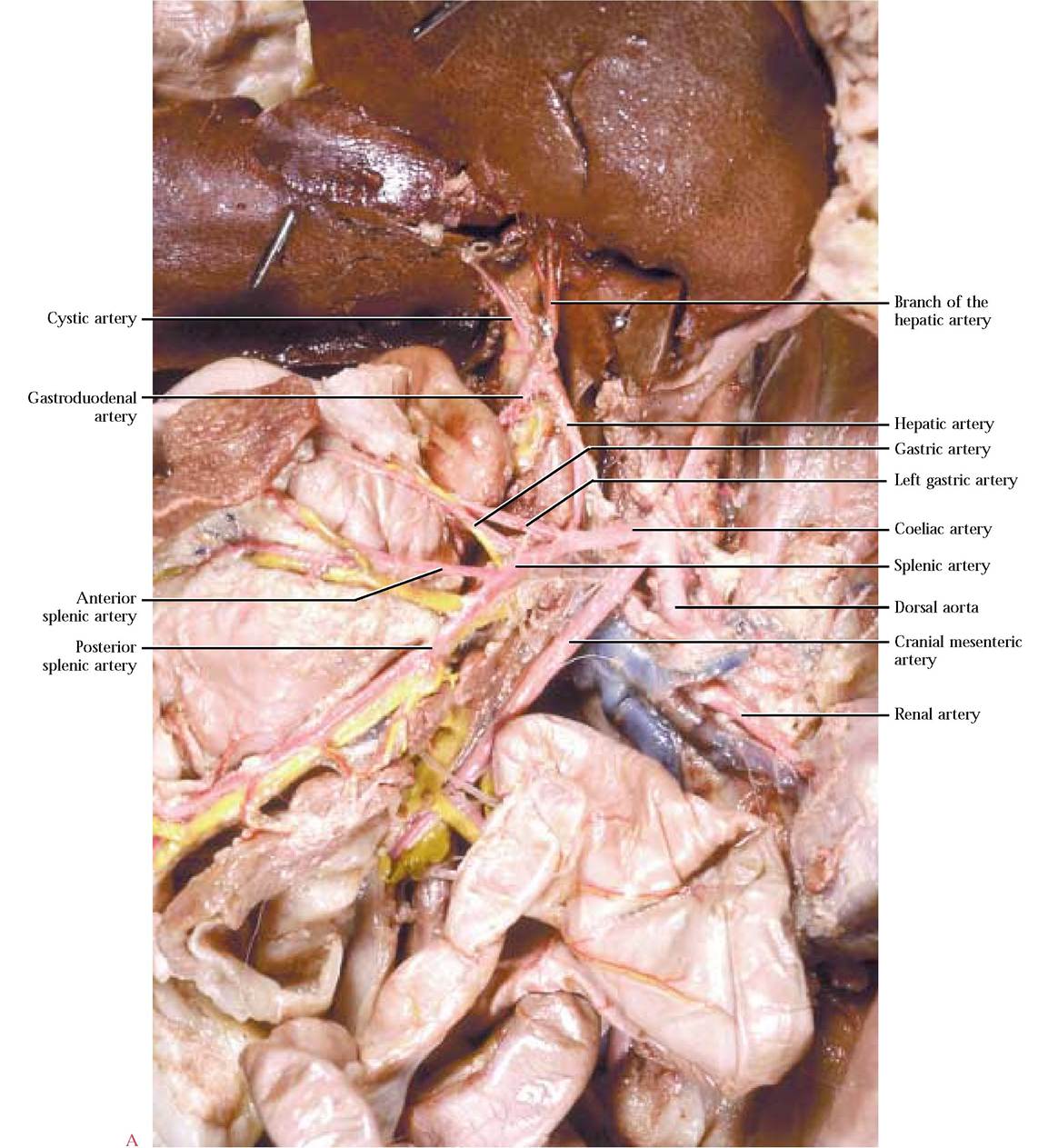
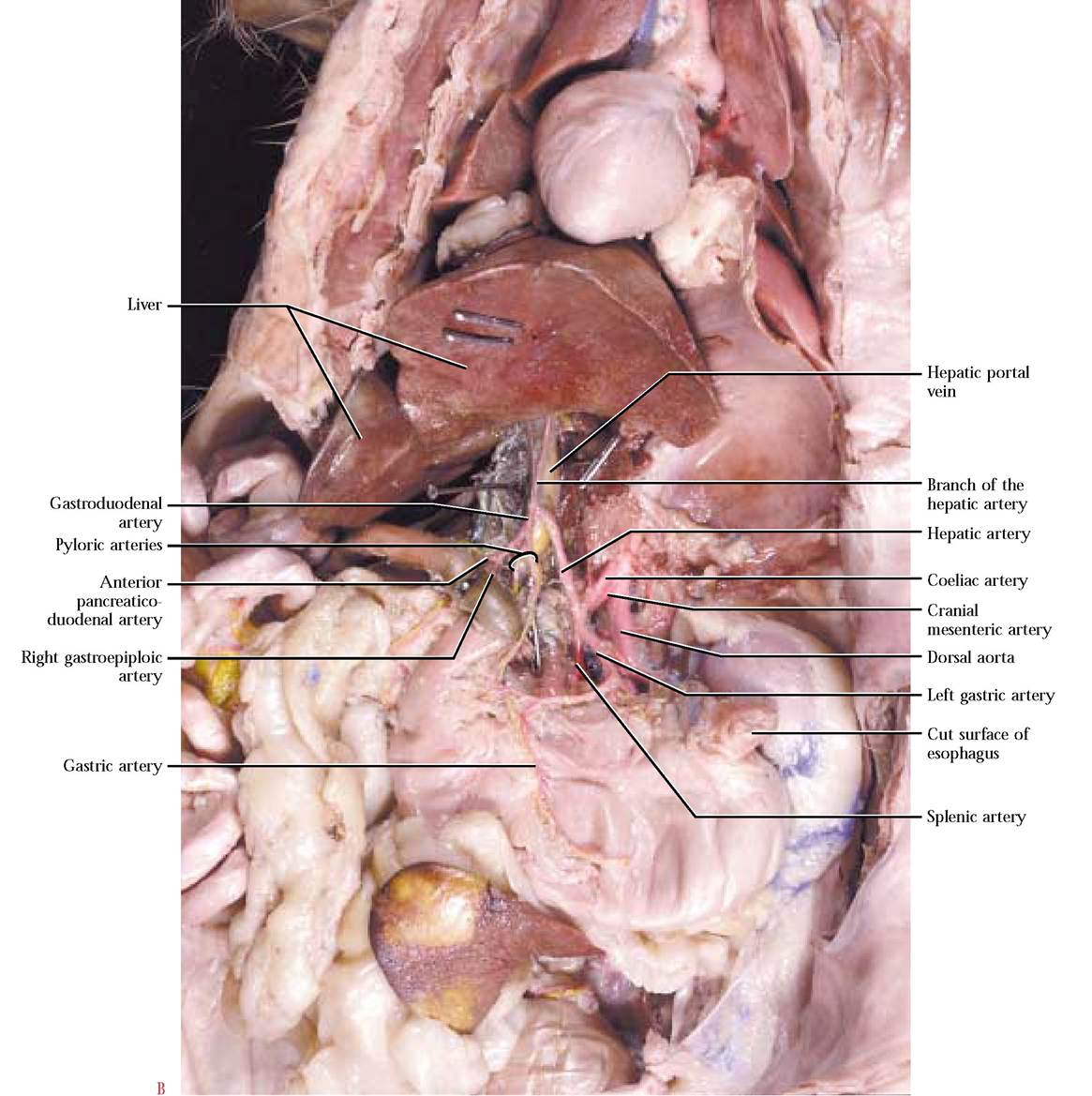
FIGURE 8-14 Coeliac artery.
The third major abdominal artery, the caudal mesenteric artery, is the smallest and least complexly branched. This vessel can be masked by the mesentery and generally must be exposed by picking away this tissue. The branches of the caudal mesenteric include the left colic artery supplying blood to the distal portion of the descending colon, and the cranial hemorrhoidal artery supplying the proximal portion of the rectum [Figure 8—13]. Notice the anastomosis that may occur between the left and middle colic arteries. The human vessel architecture in this area is similar to the cat.
Veins of the Hepatic Portal System
Drainage of the viscera, including the intestines, stomach, and spleen is accomplished by a venous architecture consisting of a specialized pattern of veins called the hepatic portal
system. A portal system is different from the common venous drainage because blood flows from a capillary bed in one organ to another capillary bed in another organ, whereas in normal venous circulation, blood is taken from the capillaries in an organ and eventually delivered to the heart. In the case of the hepatic portal system, carbohydrates and amino acids, vitamins, minerals, ions, toxins, etc. are transported from the digestive system to the liver where an astounding number of metabolic activities occur.
The hepatic portal system has capillaries on either end, preventing injection of this specialized circulation with the blue latex of the somatic veins, therefore it requires a special injection of yellow latex. As with all vessels studied and to be studied, you must carefully remove associated tissue to expose this system. Be careful not to destroy nearby vessels.
Blood is drained from the large intestine by the caudal mesenteric vein. That portion of the caudal mesenteric that drains the rectum and distal descending colon is the cranial hemorrhoidal vein while the proximal portion of the descending colon is drained by the left colic vein. The cranial mesenteric vein drains the transverse and ascending colon, cecum, small intestine and the head of the pancreas. The middle colic vein drains the transverse and often the distal portion of the ascending colon. If your cat has a right colic artery, it more than likely will also possess a right colic vein, draining the ascending colon. Drainage of the cecum and ileum is by way of the ileocolic vein. Numerous intestinal veins drain the coils of the small intestine. The posterior pancreaticoduodenal vein drains blood from the head of the pancreas and the distal portion of the duodenum. Note that the caudal mesenteric vein joins the cranial mesenteric vein [Figure 8-13 and Figure 8-15].
The stomach, the spleen and the pancreas are drained by the gastrosplenic vein. Locate the anterior splenic and posterior splenic veins draining their respective regions of the spleen. Find the left gastroepiploic vein, paralleling the artery of the same name, draining the greater curvature of the stomach and greater omentum and joining the posterior splenic vein. A variable number of gastric veins drains the body of the stomach and also join the posterior splenic vein. The pancreatic vein(s) empty into the gastrosplenic v. proximal to the junction of the cranial mesenteric v. to form the hepatic portal vein. The coronary vein draining the lesser curvature of the stomach, the right gastroepiploic vein draining the right end of the greater curvature of the stomach and anterior pancreaticoduodenal vein draining the tail of the pancreas and the proximal portion of the duodenum may join the hepatic portal v., independently, shortly after its formation. In most cats, the gastroduodenal vein, formed by the juncture of the right gastroepiploic and anterior pancreaticoduodenal veins, joins the hepatic portal vein shortly after its formation. A very small pyloric vein draining the pyloric portion of the stomach and emptying into the gastroduodenal v. may also be present. Follow the hepatic portal vein into the liver [Figure 8-16].
The hepatic portal system in humans is generally similar to that of the cat with the following exceptions. Unlike cats, the hepatic portal vein is formed by the convergence of the superior mesenteric vein (cranial mesenteric of the cat) and the splenic vein (gastrosplenic of the cat). Right and left gastric veins and a small cystic vein empty into the hepatic portal vein independently. In addition, the inferior mesenteric vein (caudal mesenteric vein of the cat), the pancreatic veins and left gastroepiploic vein merge with the splenic vein. The right gastroepiploic vein and the pancreaticoduodenal veins join the superior mesenteric vein.
Blood Vessels of the Lower Abdominal and Pelvic Regions To reiterate, the general structural plan of the circulatory system is based on vessels that deliver blood to an area (arteries) and vessels that take blood away from an area (veins). For that reason, veins of the same name almost invariably lie in close proximity to the following arteries discussed, including those arteries of both the abdominal and hindlimb regions. As you find the arteries, make sure that you find the corresponding veins at the same time.
In addition to the unpaired arteries, there are several paired vessels that typically occur at various levels along the length of the abdominal aorta. Adrenolumbar arteries arise on either side of the body a few centimeters posterior to the diaphragm and just posterior to the cranial mesenteric artery [Figure 8-17]. Their name stems from the destination of their branches. A posterior phrenic artery services the diaphragm, while an adrenal artery takes blood to the adrenal gland. The adrenolumbar a. then continues laterally to supply the dorsal body wall. The origin of both of these branches is variable, so it is possible for a number of variants to occur. Find the complementary veins, usually lying in close proximity to the artery [Figure 8-17].
A pair of renal arteries emanates from the aorta at the level of the kidneys and since the right kidney is somewhat anterior to the left, its renal artery is also slightly anterior to the left. Identify the renal veins that run parallel to the renal arteries [Figure 8-17].
As we proceed to follow the aorta caudally, the next paired arteries that are given off supply the primary sex organs of the cat. In the male, small diameter internal spermatic arteries leave the aorta and extend toward the internal inguinal ring where they accompany the vas deferens, nerves, lymphatic vessels, and spermatic veins all of which are wrapped in connective tissue known as the spermatic cord and lead to the testes [Figure 8-18]. There may be some variation in the configuration of these vessels in males. Not only may the right artery originate more posteriorly, but also the left may originate from the left renal a. In the
female, the ovarian arteries are of a larger diameter, usually originating closer to the level of the position of the ovary and then extending laterally and supplying blood to the ovary, uterine tube or oviduct and uterine horns [Figure 8-19]. An anastomosis between the branches of the uterine horns and the uterine branch of the middle hemorrhoidal artery is common.
Several pairs of lumbar arteries originate from the aorta in the lower back region, supplying blood to the dorsal muscles. The final conspicuous pair of abdominal vessels originating from the aorta just caudal to the caudal mesenteric artery is the deep ilial circumflex arteries. Each extends laterally across the iliopsoas muscles taking blood to the dorsal body wall [Figure 8-20].
Blood Vessels of the Hindlimb
At about the level of the sacrum, the aorta gives off a large external iliac artery that extends toward the hindlimb on either side of the body. In the cat, the arterial and venous pattern of the hindlimb region is very similar. However, a major exception should be noted: there is no common iliac artery, but there is a common iliac vein. In the cat, just before the external iliac artery emerges through the body wall from the body cavity, it gives off a deep femoral artery. Generally, three branches emanate from the deep femoral a., the caudal epigastric artery that extends along the abdominal surface of the rectus abdominis muscle and anastomoses with a branch of the internal mammary artery (sometimes called the cranial epigastric artery), the external pudendal artery, embedded in inguinal fat and carrying blood to the bladder and external genitalia and the third, a deep vessel, the medial femoral circumflex artery, carrying blood to the “hamstring” muscles (the biceps femoris, the semitendinosus, and the semimembranosus), as well as the adductor femoris and tenuissimus muscles[Figure 8-21]. You may notice a small branch, the external spermatic artery, joining the
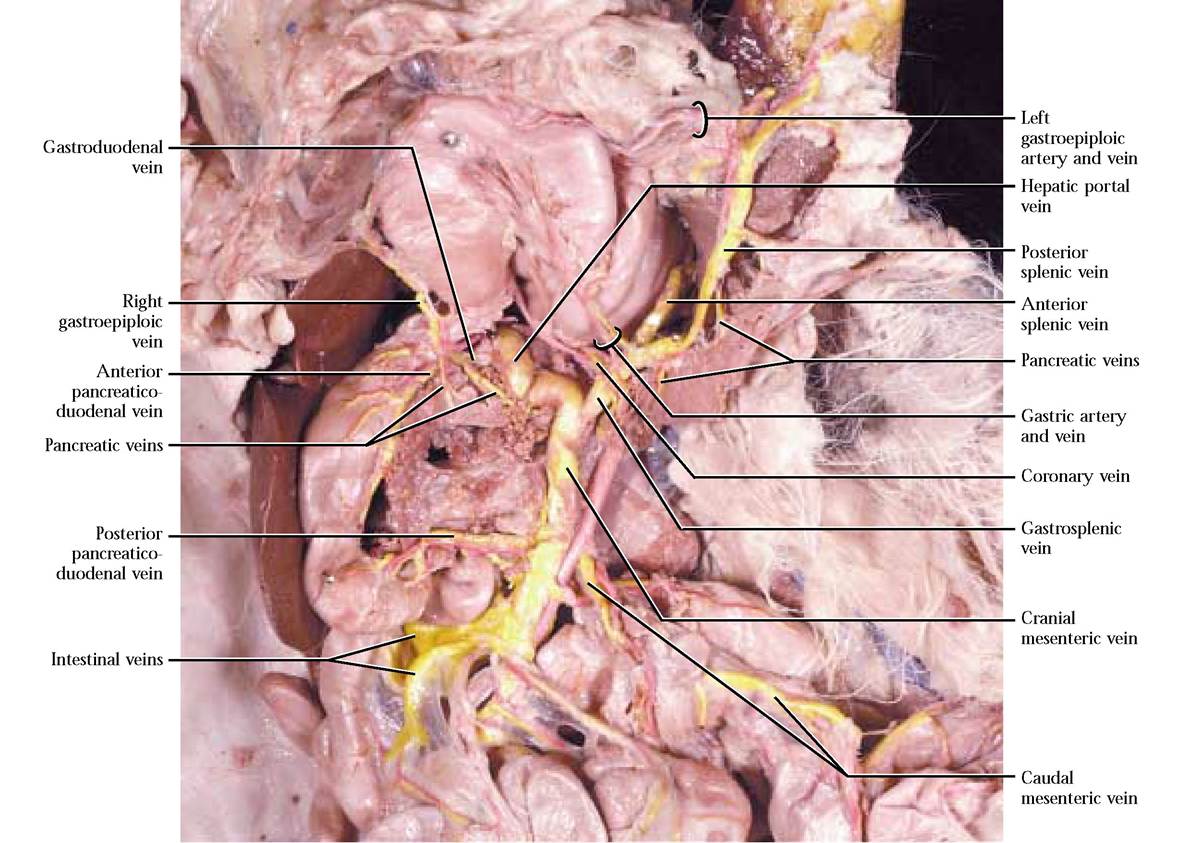
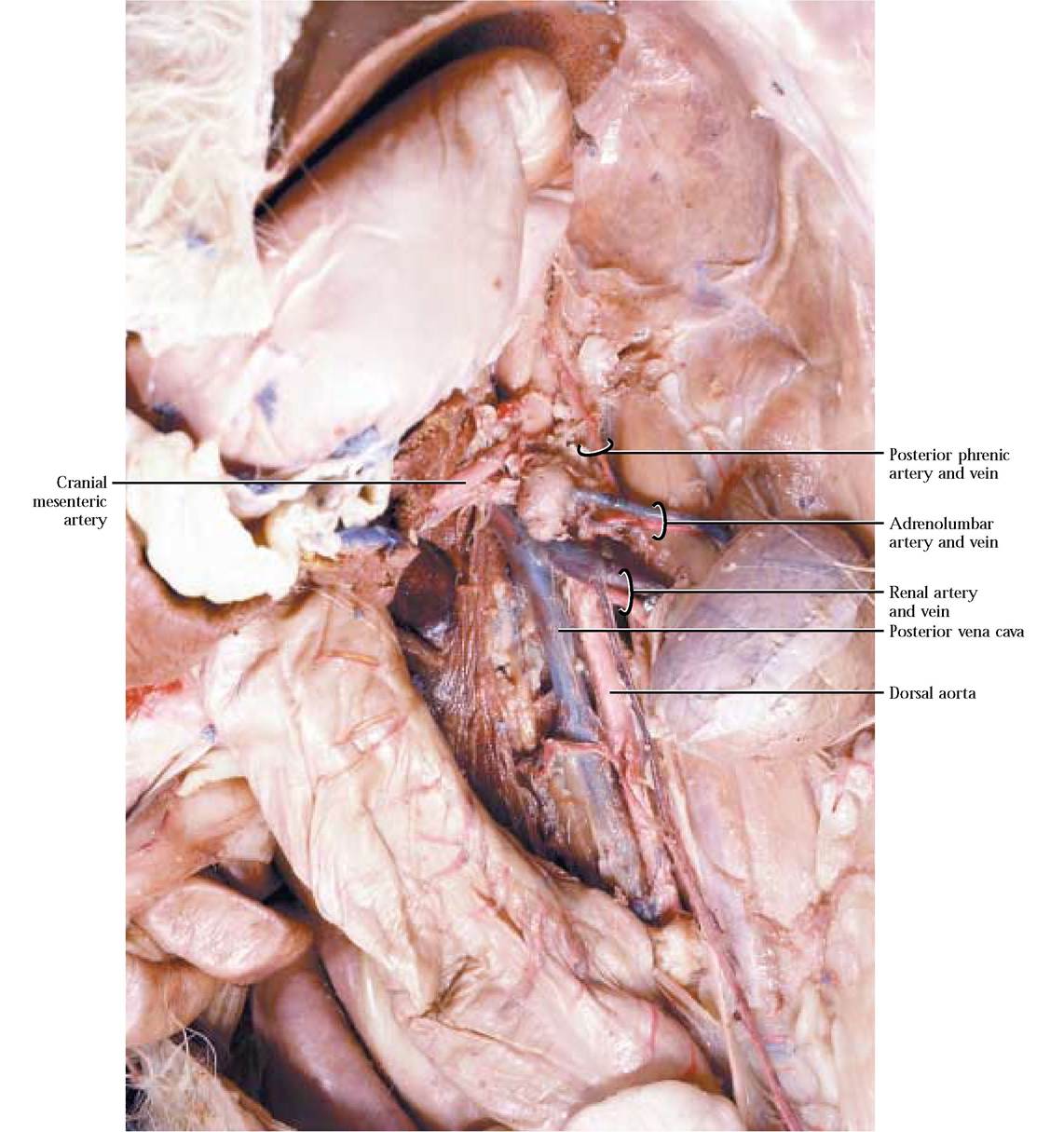
FIGURE 8-17 Renal and adrenolumbar vessels.
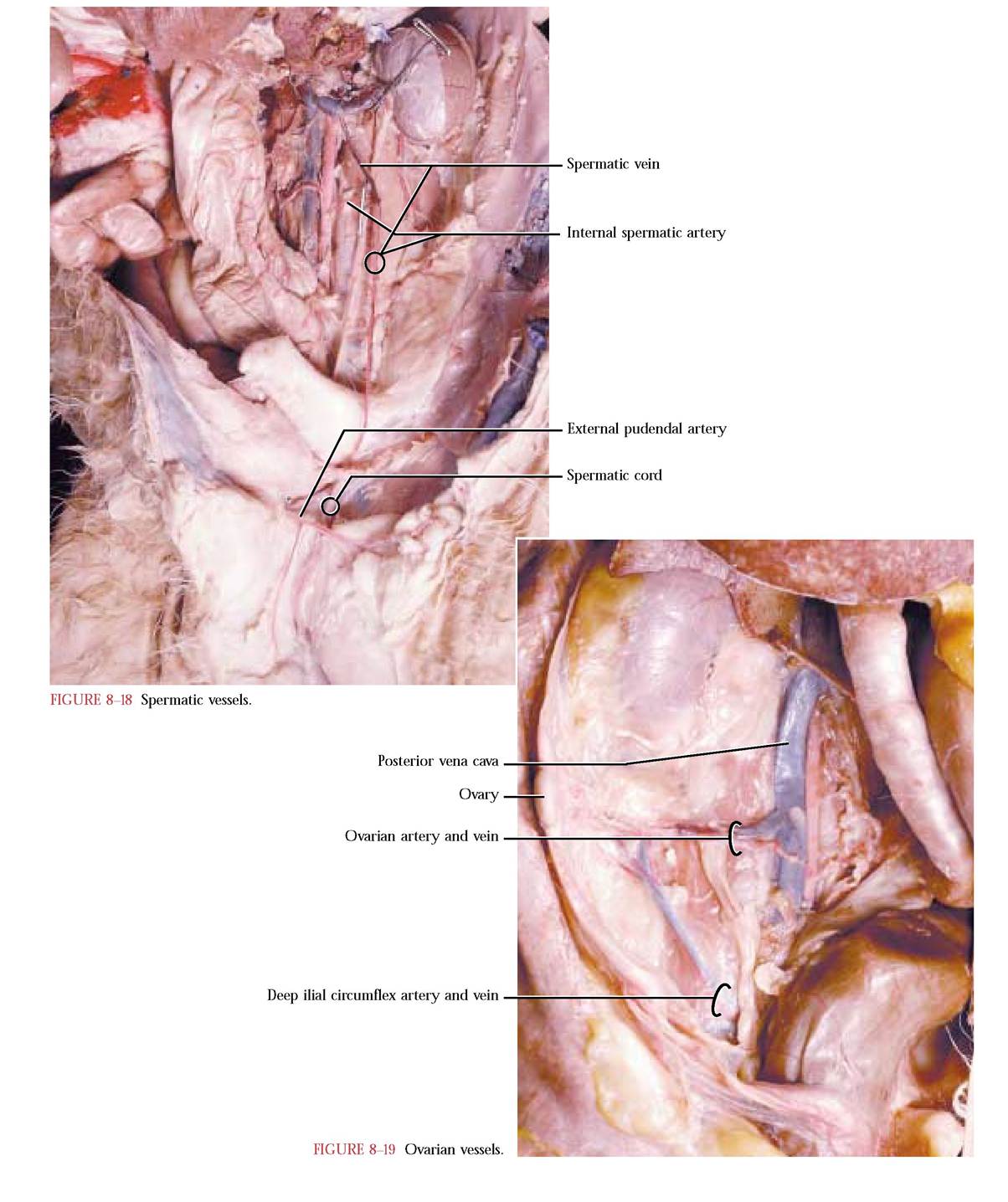
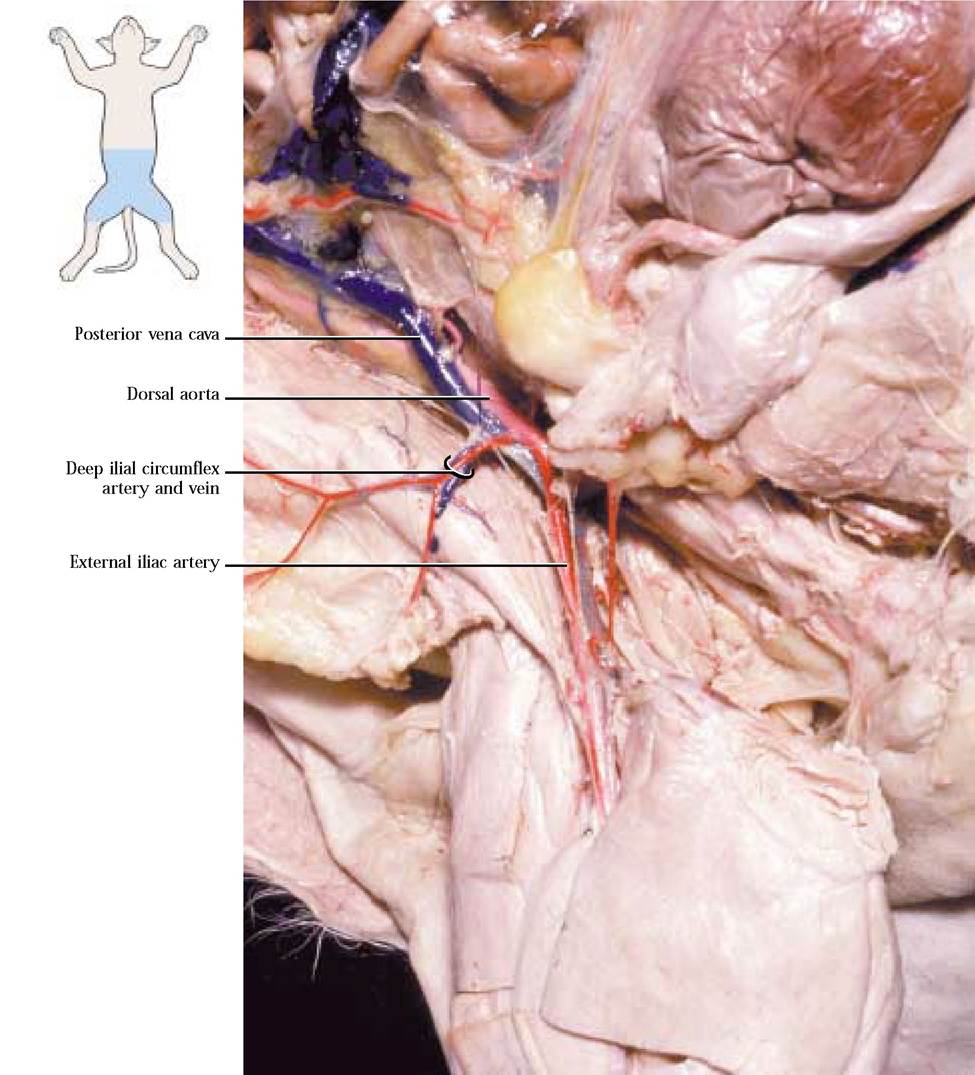
FIGURE 8-20 Deep iliocircumflex artery.
spermatic cord as it makes its journey to the scrotum in the male.
When the external iliac a. appears outside the body wall on the medial surface of the leg, it is now called the femoral artery. The first of several branches, the lateral femoral circumflex artery, emerges laterally and carries blood to the quadriceps complex (the rectus femoris, vastus lateralis, vastus medialis, and vastus intermedius) as well as the sartorius and tensor fasciae latae muscles. A somewhat more distal medial branch, the muscular artery, supplies blood to the adductors, gracilis, and semimembranosus muscles. As the femoral a. approaches the knee, it gives off the articular artery that transports blood to the gracilis, semimembranosus, and vastus lateralis muscles. At this point, or slightly distal, the superficial saphenous artery originates from the femoral and courses over the medial
surface of the thigh and shank. In the region of the knee, the femoral a. continues between the vastus medialis and semimembranosus becoming the popliteal artery. A number of branches of this artery supply blood to muscles of the shank, foot, and thigh [Figure 8-21].
Now return to the point at which the external iliac arteries originated and note that the aorta persists as a smaller version of its abdominal self and dives deeply into the dorsal portion of the pelvic cavity and is obscured by fat and connective tissue. When you dissected the reproductive system
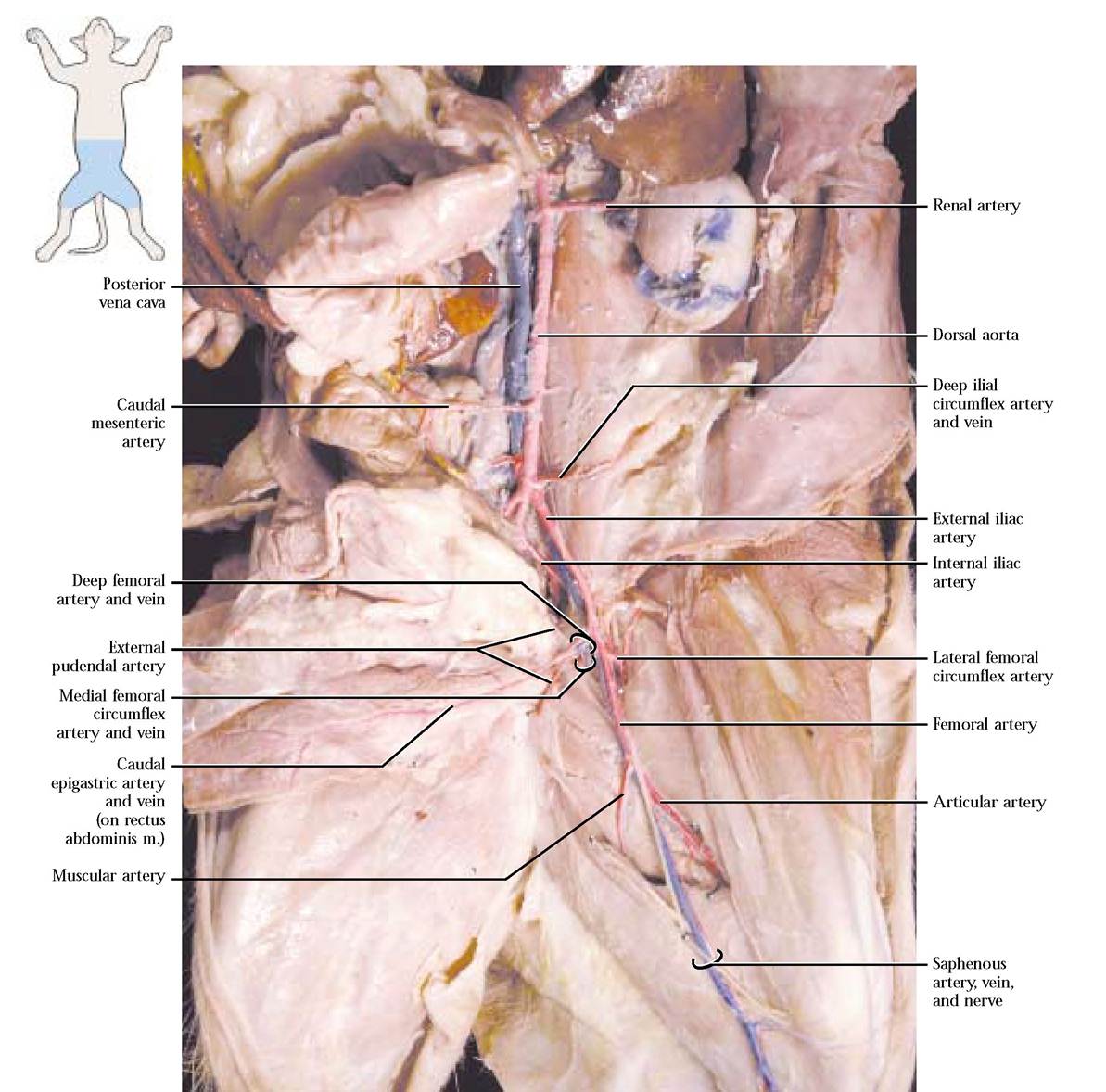
and separated the two os coxae, you undoubtedly observed some of the distal branches of this vessel. After reading the text and checking [Figure 8-21 and Figure 8-22], to observe these vessels, you will have to perform your usual careful job of slowly picking away the fat and connective tissue. When you have done so, you will see that almost immediately a pair of internal iliac arteries is given off and then the aorta persists as the small median sacral artery continuing along the dorsal aspect of the pelvic cavity and then along the ventral region of the tail as the caudal artery. The first branch of the internal iliac a. branching almost immediately is the umbilical artery extending laterally, entering the fat of the
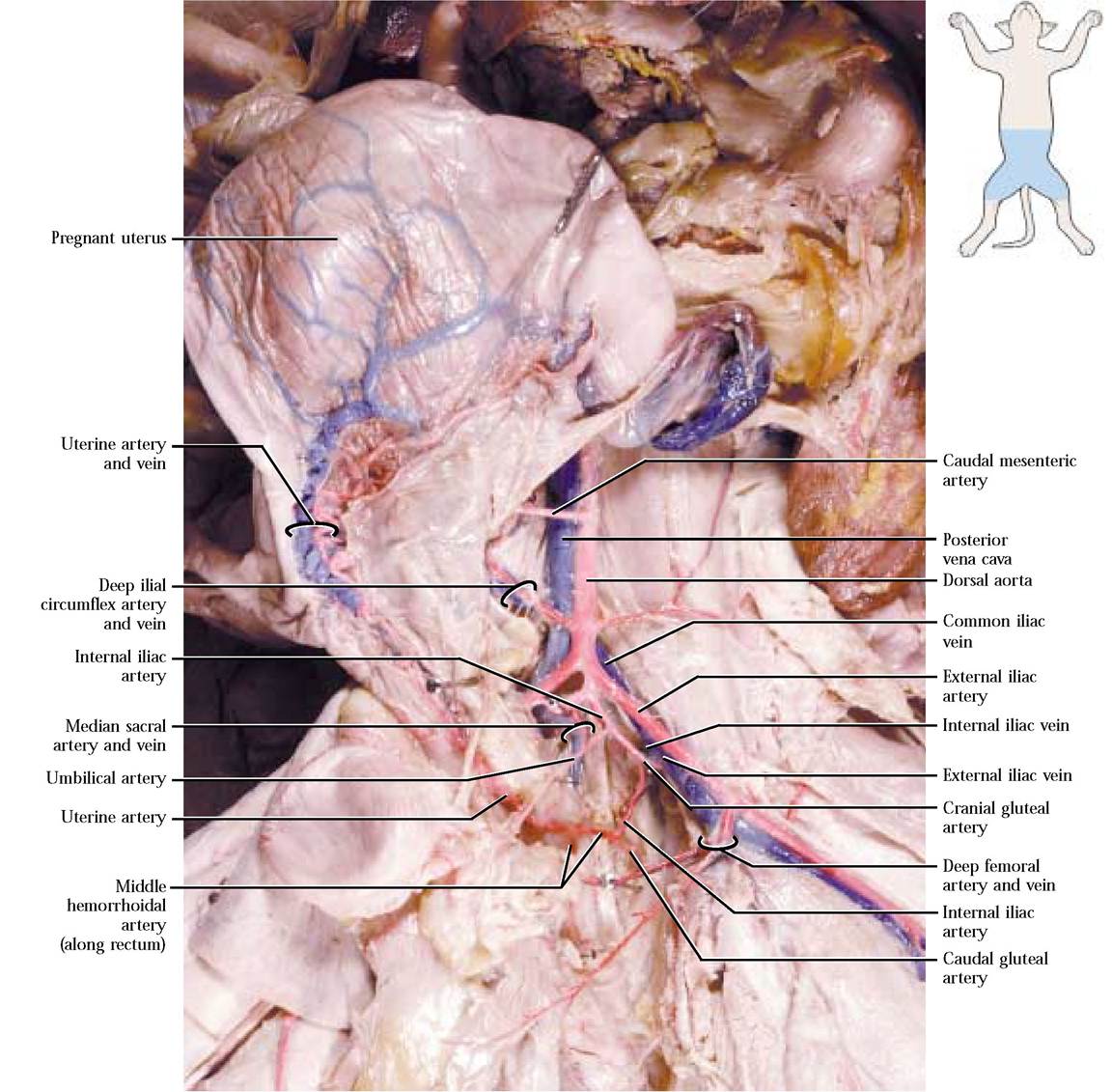
lateral vesical ligament of the bladder and supplying blood to the urinary bladder. This artery represents the remnant of the umbilical arteries that carry blood to the placenta during fetal development. The next branch is the cranial gluteal artery that turns lateral and caudal taking blood to hip muscles such as the gluteals, pyriformis, and some thigh muscles. The next vessel, the middle hemorrhoidal artery, extends medially toward various structures of the reproductive system, urinary bladder, and rectum. In females, that were pregnant or had recently been pregnant, a very conspicuous sub-branch, the uterine artery, extends cranially along the uterus and anastomoses with uterine branches of the ovarian artery. Finally, the caudal gluteal artery represents the end of the iliac artery. It supplies blood to gluteal and pyriformis muscles as well as the region at the base of the tail [Figure 8-22].
In the human, a similar pattern exists with minor modifications. A major difference is the presence of a common iliac artery that branches into the external and internal iliac arteries.
Trace the venous drainage by branches of the external and internal iliac veins and observe that they merge into the common iliac vein. The two common iliac veins join to form the posterior vena cava [Figure 8-22]. As the posterior vena cava progresses cranially, note that the veins draining adjacent organs and tissues merge with it, adding their blood to its volume. These veins should already have been identified along with the arteries of the same name. As it passes dorsal to the liver, the posterior vena cava is joined by hepatic veins draining the liver and contributing blood from the abdominal viscera that was carried by the hepatic portal vein to the liver [Figure 8-23]. To observe the hepatic veins, it is necessary to carefully scrape tissue from the cranial surface of the liver. Follow the posterior vena cava into the right atrium. The human condition is very similar.
LYMPHATIC CIRCULATION
Since it is difficult to obtain specimens with injected lymphatic vessels, the treatment of this system will be confined to a discussion. However, lymphatic nodes, glands, and organs can be identified.
The architecture of the lymphatic system consists of a network of thin walled, highly permeable vessels located throughout the body. Small fingerlike capillaries called lacteals project into the villi of the small intestine and are involved in fat absorption. The lymphatic fluid is returned to the circulatory system by way of a number of smaller vessels that coalesce into the larger thoracic duct opening into the venous system near the left subclavian vein. A large number of small organs, lymph nodes, embedded in connective tissue and often not easily identified, are clustered along the vessels. Larger lymph nodes that are more noticeable, are identifiable in the cervical, intestinal, and groin regions. As a matter of fact, the cervical nodes are often misidentified as salivary glands by students. Those found in the mesenteries of the intestines are particularly obvious and readily identified, see Figure 4-9.
Large prominent lymphoid organs that may have been previously identified during the study of other systems include the palatine tonsils, the thymus gland, and the spleen. In humans, a similar architecture exists with some modifications such as Peyer's patches, a cluster of lymph nodules associated with the intestines.
The function of this system is several-fold. At the capillarytissue level in the body, fluid pressure differences at the arteriole end cause fluids to move from the vessels into the tissues. Most of the fluid diffuses back into the circulatory system at the venule end again due to pressure differences, but a small volume remains in the tissues and is returned to the circulatory system by way of lymphatic vessels. Following fat absorption through the lacteals, these lipid molecules are transported to the blood in lymphatic vessels.
Lymph nodes and nodules are part of the internal “body sanitation and defense department. ” They are responsible for filtration and destruction of foreign proteins and particles such as viruses and bacteria. These are also the sites of lymphocyte production involved in the immune response to foreign matter.
Of the lymphoid organs, the tonsils function in a similar manner to the lymph nodes, eliminating possible foreign invaders. The thymus gland whose size is influenced by age in mammals, that is, young mammals have much larger thymuses than adults, is the site of production of lymphocytes that migrate to lymph nodes providing immune defense functions.
The largest of the lymphoid organs, the spleen is the site of immune surveillance and production of leukocytes. It is the site of aged erythrocyte destruction with release of the components into the circulatory system for production of new blood cells. A number of other organs also play an important role in the life cycle of erythrocytes. In the fetus it is a major site of erythrocyte production that generally ceases after birth, but can be reactivated in adults under conditions of erythrocyte decimation. Further, the spleen serves as a reservoir of blood platelets.
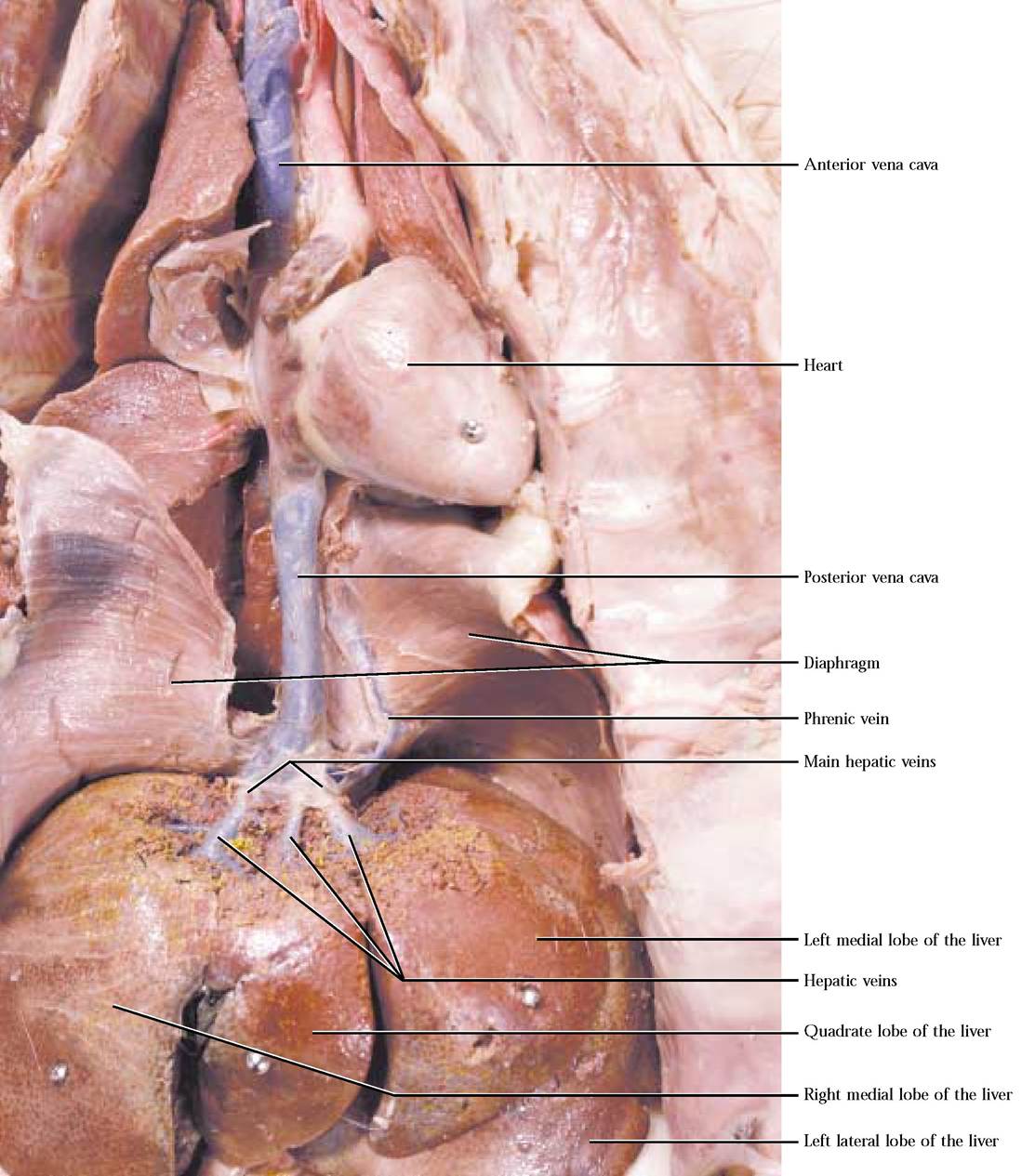
FIGURE 8-23 Circulation from the liver to the right atrium.