Cyclic AMP Is Produced by Activation of a Membrane-Bound Enzyme in Response to HormoneZNeurotransmitter Binding to Receptors
Changes in the activity of membrane-associated enzyme activities are an important mechanism of transmitting information across the cell membrane and are used by most GPCRs. Binding of a signaling molecule to receptors on the extracellular face of the plasma membrane changes the activity of an enzyme located on the cytoplasmic face.
The enzyme catalyzes a breakdown reaction; one or more of the breakdown products released into the cytoplasm are second messengers. One important such second-messenger system, and the first to have been discovered, is the hydrolytic breakdown of AΓP to 3',5,-adenosine monophosphate, or c AM P, by the enzyme adenylyl cyclase (previously called adenyl cyclase and adenylate cyclase). Cyclic AMP is the second messenger, and adenylyl cyclase is turned on or off as a result of the binding of various hormones and neurotransmitters to cell surface receptors.As summarized in Figure I-18, three distinct membrane proteins interact to produce cAMP: (1) any of several receptors, including many GPCRs; (2) the heterotrimeric G protein; and (3) the catalytic protein that actually hydrolyzes ATP to cAMP. Their interaction provides an example of the ability of biomembranes to organize biochemical reactions in space. The likelihood of three proteins colliding and thus being able to interact is much greater in the two-dimensional ‘‘phospholipid sea” than in the three-dimensional cytoplasm.
A large number of different IiormonesZneurotransmitters that bind to different membrane receptors use cAMP to transmit information across the membrane. Among the GPCRs (see '!able 1-2) and their Iiornionesziieurotraiismitters that use cAMP as their second messenger are ^adrenergic receptors that bind epinephrine or norepinephrine, increasing cAMP production and providing important regulation to almost all tissues. The starvation message carried by the binding of glucagon to its receptor (see Chapter 34) is carried to the cytoplasm by an increase in cAMP.
Vasopressin (also calledSECTIONI TheCelI
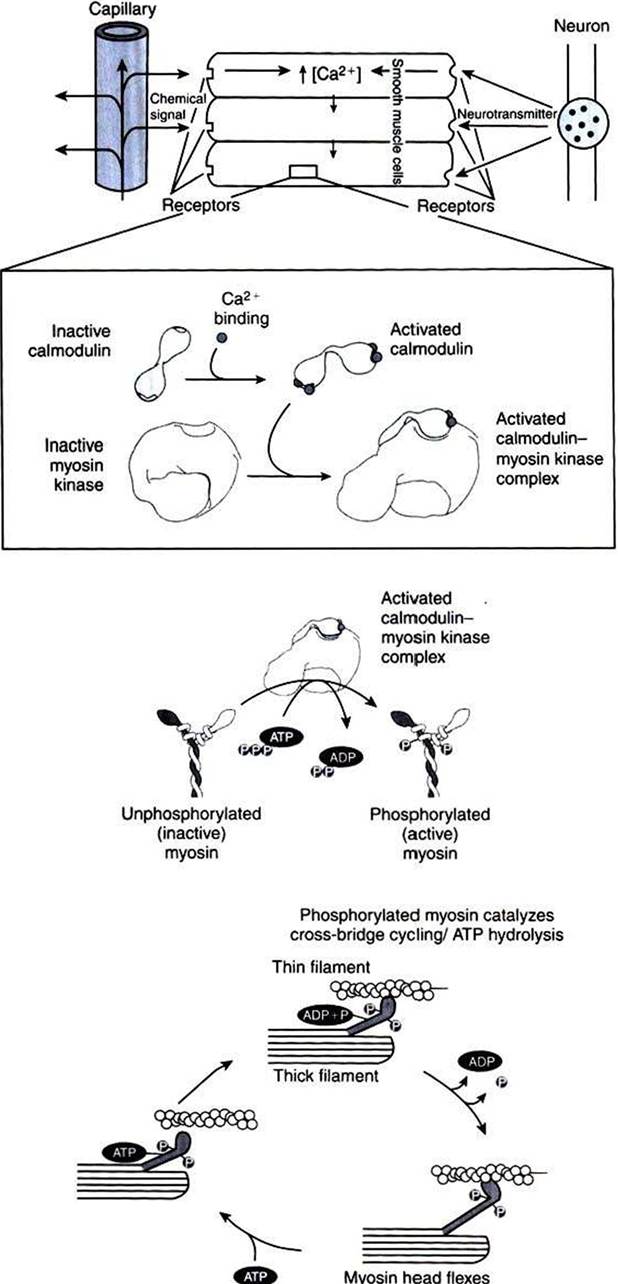
FIGURE 1-17 Role of Ca2* and calmodulin in the regulation of smooth muscle contraction. Smooth muscle regulation is more complex than regulation of striated muscle, and the account here is a simplification. Smooth muscle can be stimulated to contract by a variety of stimuli, including neural signals and soluble chemical signals, as shown here. These external signals all stimulate increased intracellular [Ca2*), which leads to smooth muscle contraction. In the presence of increased intracellular [Ca2*], the Ca2* ions bind to calmodulin, activating it by causing a conformational change. In smooth muscle cytoplasm, the activated Ca27calmodulin complex activates myosin kinase, which catalyzes the phosphorylation of myosin. Phosphorylated, activated myosin in turn catalyzes actin-dependent ATP hydrolysis (cross-bridge cycling).Thus, smooth muscle contraction is thick filament regulated, because changes in myosin activate cross-bridging, whereas striated muscle contraction is thin filament controlled, because changes in troponin and tropomyosin of the thin filament activate cross-bridging.
antidiuretic hormone, ADH) binding to its receptors in kidney cells uses cAMP to regulate urine production (see Chapter 33). A number of therapeutic drugs bind to these same receptors and mimic or prevent the physiological action of the hormone/ neurotransmitter that normally binds to the receptor.
After ligand binding, the ligand-receptor complex is able to bind to and activate the regulatory G protein (see Figure I -15, B). The G protein in turn changes shape and binds to the catalytic subunit, altering its shape and regulating its ability to bind ATP, and hydrolyzes the catalytic subunit to cAMP (see Figure 1-18). There are two types of G proteins in the adenylyl cyclase system, which differ in their Ot subunit.
The Gs (more specifically, Gαs, s for stimulatory) activates the catalytic subunit; this is the G protein shown in Figure 1-18. A different G protein, the α subunit of Gi, inhibits adenylyl cyclase when activated. Some diseases are the result of the binding of bacterial toxins to the G proteins. Cholera symptoms result in part from the binding of the toxin of the bacteria Vibrio cholerae to the Gs protein, and the irreversible activation of the Gs protein, which in turn irreversibly activates the catalytic subunit. Pertussis (whooping cough) toxin binds irreversibly to and activates Gi, thus inactivating the enzymatic activity.As suggested by the inhibitory G protein (Gi), regulated decreases in cAMP concentrations are an important part of the cAMP second-messenger system. There are two mechanisms for such decreases: decreasing the rate of cAMP production and eliminating cAMP after formation. The former is achieved by Gi inhibiting the catalytic subunit. Certain inhibitory receptors specifically interact with Gi. Opium and drugs derived from it, such as codeine and morphine, are examples of signaling molecules that bind to inhibitory GPCR (opioid) receptors, activate Ci1, and inhibit production of cAMP. Other examples are norepinephrine and epinephrine acting through α2-adrenergic receptors. Recall that these same neurotransmitters activate adenylyl cyclase when bound to β- adrenergic receptors. This is another example of the principle that the receptor/ligand complex contains the information, not the IiormoneZneurotransmitter itself.
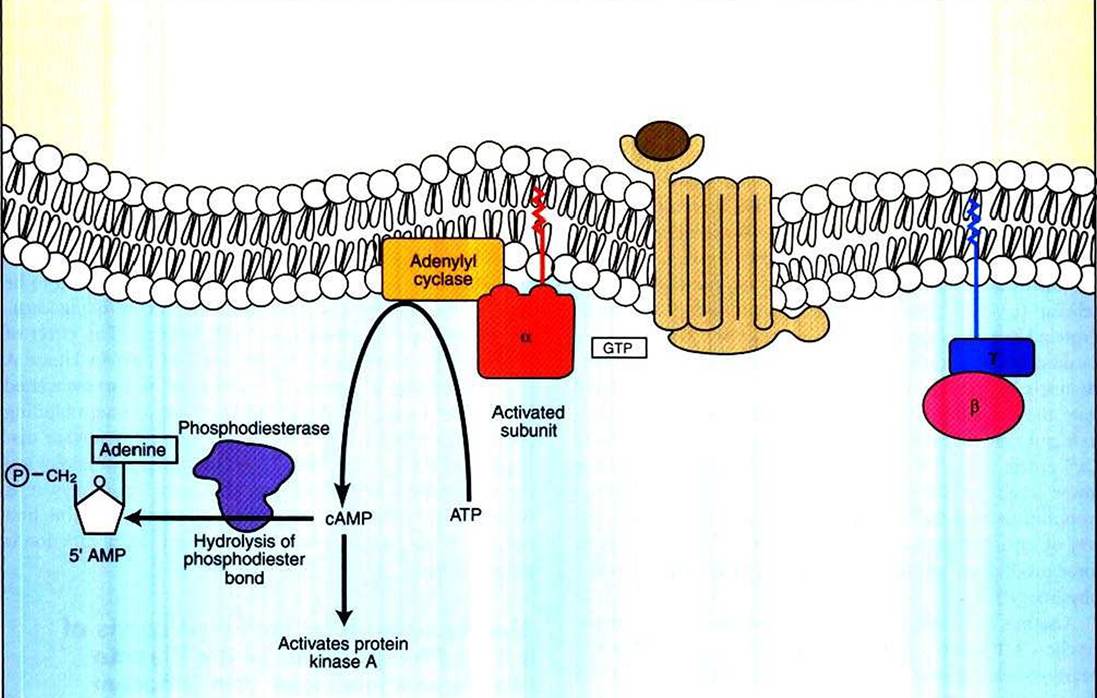
FIGURE 1-18 Activity of cyclic adenosine monophosphate (cAMP) as a second messenger. Cyclic AMP is generated through GPCR-Iinked activation of adenylyl cyclase, causing hydrolysis of ATP to cAMP.
The cAMP thus generated binds to and activates a specific protein kinase, protein kinase A, which in turn can phosphorylate and change the activity of various cellular substrates. Once generated, cAMP is broken down by phosphodiesterase (PDE)t which hydrolyzes cAMP to "normal" adenosine monophosphate (i.e., 5' AMP).The other control on cAMP levels is elimination of cAMP after formation. This is regulated by the enzyme cyclic nucleotide phosphodiesterase. This enzyme hydrolyzes the 3' ester bond of the phosphate to the sugar to produce “plain” 5z AMP (see Figure 1-18). As with myosin kinase discussed earlier, phosphodiesterase is a Ca2-*/calmodulin-activated enzyme, so in many cells the activities of the Ca2* and cAMP second- messenger systems antagonize one another.
The increase or decrease in cAMP concentrations most often affects cell function through cAMP’s interaction with a particular protein kinase. This protein kinase is called cΛMP- dependent protein kinase, or protein kinase A. This protein kinase is distinct from the Ca2’/calmodulin-dependent protein kinase discussed earlier, although the basic outline of action is similar. Protein kinase A is activated by binding cAMP. The higher the concentration of cAMP in a cell, the greater is the number of active protein kinase A molecules. The activated kinase binds to proteins and ATP, hydrolyzing the ATP and phosphorylating the protein. As shown in previous examples, this phosphorylation alters the activity of the target protein, altering its particular characteristic function: catalysis, transport, coupling, and so forth.
Mammals respond to a stressful stimulus by increasing the force and rate of heart contraction, among other physiological effects. This increase in force demonstrates the role of cAMP as a second messenger and the role of Ca2* in GPCR signaling, and it is another example of physiological Rube Goldberg devices based on allosteric changes in proteins.
The stressful stimulus causes the adrenal medulla to release epinephrine to the blood, and sympathetic nerves release norepinephrine to the heart. Both catecholamines bind to β-adrencrgic GPCRs on the cardiac muscle cells. The receptor-ligand interaction stimulates adenylyl cyclase by way of Gs, increasing intracellular [cAMP], thus increasing protein kinase A activity. Protein kinase A phosphorylates a number of substrates in the cardiac muscle cells, including voltage-dependent Ca2* channels in the plasma membrane. In the phosphorylated state, these channels remain open somewhat longer in response to membrane potentials above threshold. Consequently, more Ca2* enters the cell for a given electrical stimulation than at lower levels of cAMP. The increase in Ca2+ allows ∏1OΓC troponin to bind Cr+: Π10Γθ tropomyosin moves out of the way of myosin heads, causing more cross-bridging and more force production. (Rube Cioldberg would have loved modern physiology!)Another cyclic nucleotide, cyclic guanosine monophosphate (cyclic GMP), also serves as a second messenger but is not nearly as widely used as cyclic AMP. Cyclic GMP (cGMP) is the second messenger stimulated by opsins (see Table 1-2) in the rod cells of the retina underlying vision and also causes relaxation of some vascular smooth muscle, including that responsible for penile erection (i.e., blood flow into the corpus cavernosum). The role of cGMP in erections is mediated by its activation of cGMP protein kinases, similar to cAMP action via protein kinase A. Adivation of cGMP-dependent protein kinase causes relaxation of certain smooth muscles, including those responsible for blood flow to the corpus cavernosum. This has an important clinical correlate: the drug Viagra (sildenafil) inhibits the breakdown of cGMP by a cyclic nucleotide phosphodiesterase, thus increasing blood flow to the penis, and aids erection, but only if neural signals (i.e., sexual stimulation) have stimulated cGMP production initially.
This is a good example of how the multistep pathway of cell signaling provides multiple potential sites for appropriate therapeutic intervention; a drug that simply stimulated cGMP production would cause inappropriate erections, whereas inhibiting its breakdown aids timely erections. Although used mostly by men, sildenafil is also occasionally used for stallions to assist them in “covering” a mare.In addition to activating protein kinases, cAMP and cGMP can also bind directly to and cause opening of a class of ligand-gated ion channels, cyclic nucleotide-gated ion channels. These ion channels are atypical in that their structure resembles voltage-gated K, channels, but they open by directly binding a cyclic nucleotide. These channels play an important role in smell, for which cAMP is the relevant second messenger. In vision, as noted earlier, cGMP is the second messenger, and mutations in the cyclic nucleotide-gated ion channels of cones are responsible for most forms of complete color blindness (but which is rare).
The examples of physiological control by second messengers discussed thus far are short time-scale changes (seconds to hours), which historically have been the purview of “physiologists.” It has become increasingly clear, however, that most, if not all, major signals have longer-term (days and weeks) effects based on changes of gene transcription, which in turn mediates changes in growth, differentiation, and longterm behavior. For example, cyclic AMP is now known to be an important regulator of gene transcription controlling learning, production of gametes, and cell division. The effect of cAMP on gene expression is the result of protein kinase A phosphorylating a specific transcription factor associated with cAMP signaling (“cyclic AMP response element binding protein,” or CREB). While space does not allow further discussion of the transcriptional roles of “classic” physiological signal pathways, when dealing with signal pathways it is worthwhile to keep in mind the disclaimer in the first paragraph: only a highly simplified account of cell function is presented here!