The Receptor-Mediated Hydrolysis of a Rare Phospholipid of the Plasma Membrane Produces Two Different Second Messengers with Different Actions
Anoiher second-messenger system differs from both Ca2' and cAMP in that r∣∣,o distinct second-messenger molecules are produced as a result of an CfiZy∏]il∏C IlChVfltioil by a single receptor/ligand complex.
Phosphatidylinositol (PI) is a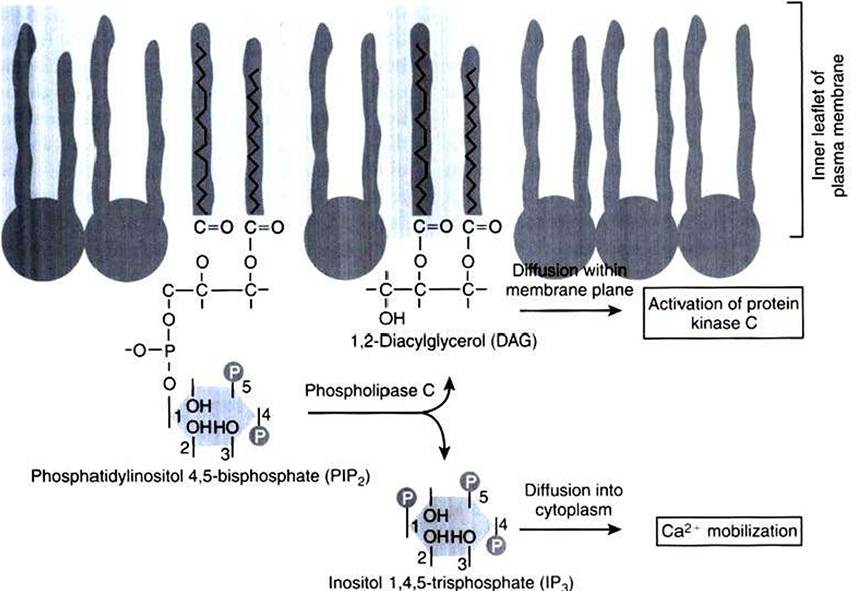
FIGURE 1-19 Hydrolysis of a membrane lipid to produce two second messengers. After appropriate receptor and G-protein activation, the rare membrane phospholipid shown to the left, phosphatidylinositol 4,5-bisphosphate (PIP2), is hydrolyzed into two separate second messengers by phospholipase C.The phosphate "head" of the PIP2 molecule is cleaved to produce the soluble messenger inositol 1,4,5-trisphosphate (IP3), which mobilizes intracellular Ca2*, as well as the Iipidic messenger diacylglycerol (DAG), which remains in the membrane and activates protein kinase C.
membrane phospholipid that can accept additional phosphate groups by reaction with the —OH groups on the inositol (Figure 1-19). Phosphatidylinositol 4,5-bisphosphate (PIP2) is the membrane phospholipid that is broken down to produce two important second messengers. PIP2 is hydrolyzed to diacylglycerol (DAG) and inositol 1,4,5-trisphosphate (IPJ by a receptor-mediated enzyme called phospholipase C (PLC) or phosphoinositidase. Although many distinct processes are controlled through the PIP2 path, it plays a particularly important role in control of growth and of receptor-mediated secretion. The effect of acetylcholine acting through muscarinic receptors (not the nicotinic receptor/ion channel of the nerve-muscle synapse) is often transmitted and transduced through activation of the PIP2 pathway.
The events involved in the receptor-mediated production of IP3 and DAG from PIP2 are similar to those in the production of cAM P.
The membrane system appears to consist of three distinct intrinsic membrane proteins: (1) any of several different GPCRs, including the muscarinic acetylcholine receptor and the receptors for some growth factors; (2) a heterotrimeric G protein, similar but not identical to Gs of the cAMP pathway; and (3) the hydrolytic enzyme PLC. A hormone/neurotransmitter or growth factor binds to the receptor, forming a receptor/ligand complex. This complex activates the G protein, which in turn activates the hydrolytic enzyme. At present, only a stimulatory G activity on PLC is known; there is no evidence for an inhibitory G activity in this system.The activation of the hydrolytic enzyme increases the concentration of IP3, which is water soluble and thus diffuses through the cytoplasm. IPκ binds to and opens ligandgated Ca2 ‘ channels in the endoplasmic reticulum. This releases Ca2' from that high [Ca2J compartment into the cytoplasm. Ca2 thus becomes the “third messenger” in this system (although this term is not in widespread use) and is another example of a role of Ca2' in GPCR signaling. The ensuing increase in cytoplasmic ∣Ca2*∣ affects cellular function by the same mechanisms outlined earlier for Ca2* as a second messenger (e.g., binding to calmodulin), with the Ca2'/calmodulin complex in turn activating various enzyme activities. In receptor-mediated secretion, for example, the binding of acetylcholine to muscarinic receptors in the pancreas (the organ that secretes digestive enzymes) causes an increase in PIP2 breakdown and an increase in cytoplasmic IPv The IP; opens ligand-gated Ca2, channels in the endoplasmic reticulum, and intracellular [Ca2+∣ increases. The process then becomes similar to smooth muscle contraction. Calmodulin binds Ca2∖ and the complex activates a protein kinase.
However» rather than activating myosin, as for smooth muscle, activation of this protein kinase causes exocytosis of secretory vesicles (membrane bubbles full of secretory product) with the plasma membrane, releasing the enzymes into an extracellular space that is contiguous with the gut.DAG is also produced on activation of PLC, but it is not at all soluble. DAG diffuses in the plasma membrane, binding to and activating membrane-associated protein kinase, protein kinase C (PKC). PKC is not an intrinsic membrane protein and can bind reversibly to the cytoplasmic face of the plasma membrane. PKC phosphorylates other proteins and changes their activity. Because of the membrane-bound character of the enzyme, most evidence indicates that PKC phosphorylates membrane proteins such as receptors and ion channels, regulating their function. In the case of the secretory response to some hormone/neurotransmitter stimulus, PKC generally acts separately but additively with IP5 to produce the response. As with cAMP and protein kinase A, however, much interest focuses on longer-term effects of DAG activation of PKC, particularly its role in growth control and cancer. A class of chemicals long known to promote the onset of cancer, phorbol esters, is a potent substitute for DAG at activating PKC. It is now known that PKC indirectly activates an important transcription factor involved in cell proliferation, nuclear factor kappa B (NF- κB). Thus, as a second messenger and as with cAMP» DAG has both short-term effects and longer-term transcriptional effects.