Death by Cancer Is Usually the Result of Its Spread, Not the Original Tumor
Death from cancer is often the result of the spread of the cancer from the initial tumor, the primary tumor, to various distant sites. This process of cancer cells colonizing other tissues is called metastasis.
For some cancer types, including leukemias, and those of the brain, the primary tumor itself can be fatal. In contrast, the primary tumor for melanoma, a very deadly cancer, is little more than a mole on the skin that does not become life threatening until these cancer cells spread. Although metastasis is the deadliest aspect of cancer, much less is known about it than about the dysfunctions of cell growth leading to the primary tumor.The best understood aspect of metastasis is that it occurs by a multistep process called the metastatic cascade. In this step-by-step process, cells escape from the primary tumor, breaking through tissue barriers to gain access to the circulatory system. The cells are carried until they escape the circulatory system to invade a new tissue (Figure 2-13). The steps of the metastatic cascade suggest that dysfunction of three broad types of cellular function are particularly important:
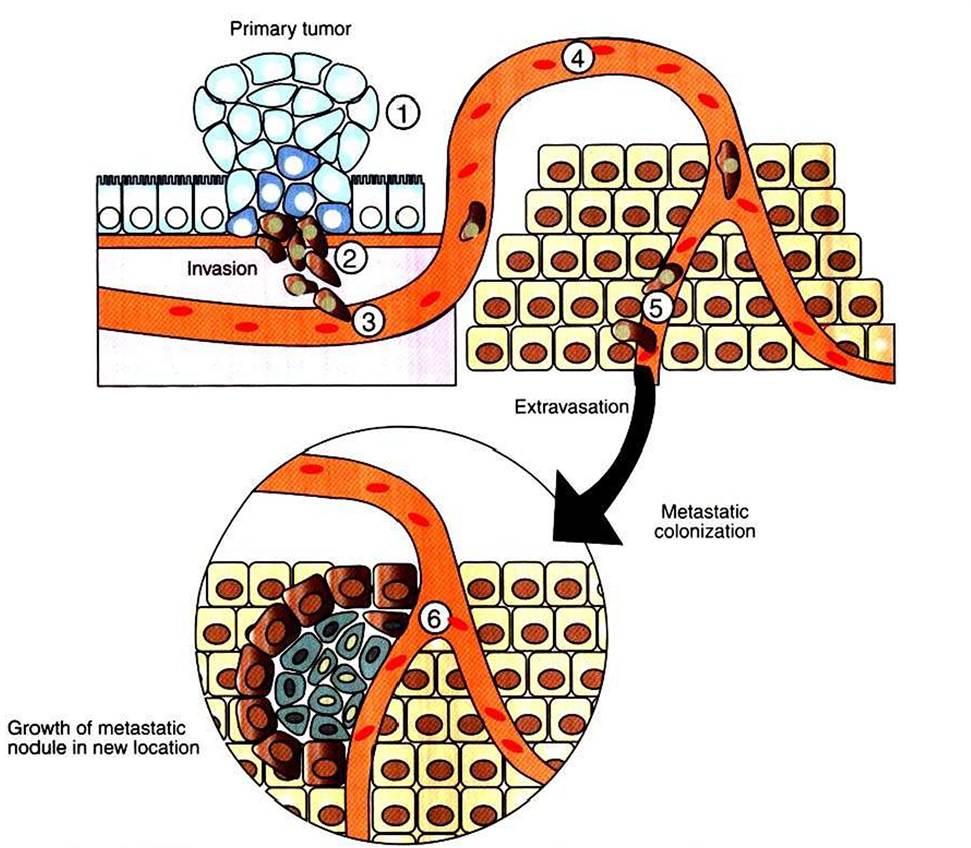
FIGURE 2-13 Metastatic cascade, the path from primary tumor to metastatic tumor. Cells of the primary tumor alter their cell adhesion and motility properties to migrate away from the primary tumor site (U. These cells secrete proteases to digest their way through the surrounding tissue (2). They then crawl into the vasculature (3), a process called intravasation, where they are then carried passively around the circulation (4). At some point, they adhere to the sides of the blood vessel and crawl out of the vasculature (5), a process called extravasation. Some metastatic cells are able to colonize the new location to form a new, deadly metastatic tumor (6).
cellular adhesion, cellular motility, and secretion of proteases. How these dysfunctions arise from the genetic damage of growth in the primary tumor is, once again, unknown, but mutation resulting from the genetic instability of the primary tumor is typically suggested as a link.
The first step of the metastatic cascade is the loss of cell adhesion by the cancer cell, both to neighboring cells and to the extracellular matrix (ECM). Accordingly, many types of cancer cells show greatly reduced expression of a cell-cell adhesion receptor, E-cadherin, important for epithelial adhesion. Similarly, primary tumor cells show a wide variety of abnormalities in the number and type of cell-ECM adhesion receptors, integrins, they express. In addition to loosening the bonds to the primary tumor, allowing cells to escape, one hypothesis is that these changes in cell adhesion molecules underlie the curious tendency of various cancers to metastasize preferentially to certain other tissues. Melanoma, for example, has a strong tendency to metastasize to the brain and to bone. Melanoma’s particular array of abnormal (for skin) adhesion molecules may represent a “postal code” favoring delivery to a particular distant site.
Having altered its adhesion, enabling escape from the primary tumor, the metastatic cell must make its way toward the circulatory system, enter the circulation (called intravasation) to “hitch a ride” around the body. Although “circulation” typically refers to the bloodstream, cancer cells can also be disseminated by traveling within the lymphatic system, which collects extracellular tissue fluid for return to the blood. Indeed, invasion of lymph nodes, which are major collection sites for extracellular fluid and debris, is a common test for initial metastases. For either route, however, the cells ability to achieve intravasation depends on altering normal motility and expressing proteases. Most animal cell types are capable of “crawling” locomotion using actin and myosin mechanisms similar to muscle contraction (see Figure 1-4).
This crawling locomotion is similar to the motility of amebae. Migrating breast cancer cells have been imaged directly and show solitary cells with amoeboid morphology. The entire actin and myosin system of most cancer cells is dysregulated, causing changes in cell shape and the ability and tendency to Iocomote. For example, normal skin cells are generally quite stationary, but melanoma cells are highly motile. The dysregulation of the actomyosin system results in part from mutations of the Rho family of small, Ras-Iike G proteins, mentioned briefly earlier. Mutations in rho are common among highly metastatic melanoma cells, but such mutations are rare among weakly metastatic melanoma cell lines.Because cells in solid tissues are crowded together, increased motility appears to be helped by secretion of proteases that digest some of the cell matrix “obstacles” in the cancer cell's path. Epithelial cells give rise to approximately 80% of human cancers. As noted earlier, all epithelial cells are attached to an ECM, which is characterized by a particular type of collagen. Proteases specific for (his type of collagen are generally overexpressed by metastatic cells. The number of different proteases and the net amount of protease secreted tend to increase with increasing metastatic potential. In addition, cancer cells appear to have the capacity to cause surrounding normal cells to increase their secretion of proteases. Proteases not only aid the metastatic cell in intravasation, but also stimulate cell survival and proliferation by largely unknown mechanisms. Flowever, not all ECM represents an obstacle to movement that must be ρroteolytically degraded. Some types of ECM appear to provide stimulatory pathways for the migrating cells to follow toward the circulation. Migration toward the circulation is also aided by chemoattraction; epidermal growth factor in blood vessels seems to be an attractant for breast cancer cells.
After intravasation, the metastatic cell rides around the circulation until it can attach to the vessel wall.
Then, reversing the process of intravasation, the cell “climbs out” of the circulation, called extravasation. As one might expect, this also depends on changes in adhesion, motility, and protease secretion. After extravasation, the metastatic cell must be able to survive and proliferate in its new environment.Fewer than 1 in 10,000 cells escaping the primary tumor colonize a new location successfully. Il was once assumed that this high rate of failure reflected an “exceedingly rough ride” around the circulatory system. More recent evidence suggests that the limiting factor of metastasis is the survival of the cell in its new location. This represents another example of the natural selection, the “microevolution,” occurring in cancer. The foreign environment exerts a strong negative selective pressure on cancer cell arrivals, and the vast majority do not survive. Ironically, our current thinking about metastasis is similar to the “seed and soil hypothesis” first proposed in 1889. Metastasis requires a cancer cell from the primary tumor (the seed) capable of carrying out (selected for) all the steps of the metastatic cascade, and the metastatic cell must colonize a region (the soil) appropriate for its subsequent growth. Normal cells cannot survive in a new location within the same body. Different tissues have different chemical milieus (e.g., different mixes of growth factors), and these are specialized for the survival and growth of the particular cell types found in the region. The process of metastasis necessarily selects for cells capable of growing in a foreign environment. Il is thought that most metastatic cells become dormant in their new location and that additional genetic changes are required and must be selected to enable uncontrolled growth. Fewer than 1% Ofmicrometastases grow to a clinically relevant size. ∕∖ key aspect of this selection process for uncontrolled growth in the foreign location is the secretion by the cancer cells and by surrounding cells of a variety of mediators to stimulate growth of new blood vessels to supply the tumor.
Another key feature of selection is remodeling of the local ECM, which normally is required for proliferation. These phenomena of inducing blood supply and remodeling of the ECM are other aspects of metastasis that are poorly understood.Indeed, it is poorly understood how metastasis actually leads to death, except that it often involves a profound and overall wasting of the body, cachexia. Cachexia is complex, but it is caused in part by generalized inflammatory reactions, which attack other aspects of the patient's physiology. Thus, it appears that the presence of foreign cells, selected for growth in an abnormal location, causes the body’s defense mechanisms to be fully mobilized and attack itself, a nightmare of biological “friendly fire.” Presumably, the “foreignness” of the metastatic tumor explains the highly inappropriate response, which primary tumors typically do not instigate. For this reason, in part, complete removal of the primary tumor before metastasis occurs often leads to total recovery. In other J
cases, death from metastatic disease, similar to death from some primary tumors, is the result of cancer cells simply overwhelming a vital organ, leading to organ failure.