HAIR
Hair is a mammalian feature, diagnostic of the class. In most species a thick haircoat is spread over the body, except about the mouth and other openings and on the surfaces of the feet; in a few, including the domestic pig (though not its ancestors), the covering is sparse (Figure 10-10, E).
The individual hairs take a variety of intergrading forms, but only three need be distinguished here: straight, rather stiff guard hairs provide a “topcoat”; fine, wavy wool hairs provide an “undercoat”; and stout tactile hairs of restricted distribution are associated with touch receptors.Guard hairs mostly lie close against the skin and sweep uniformly in broad tracts, giving the coat a smooth appearance disturbed only by the whorls, crests, and partings formed where different streams converge and combine or diverge from one another. The regularity of the arrangement is significant because it promotes the runoff of rain, preventing the chilling that would occur if water were allowed to penetrate the pile to reach the skin. Occasionally, animals are born with a disturbed coat pattern, which may seriously impair their ability to withstand severe weather. However, as with so many other features, breeders have chosen to promote deviant mutant arrangements as attributes of particular breeds, particularly of dogs, cats, and rabbits.
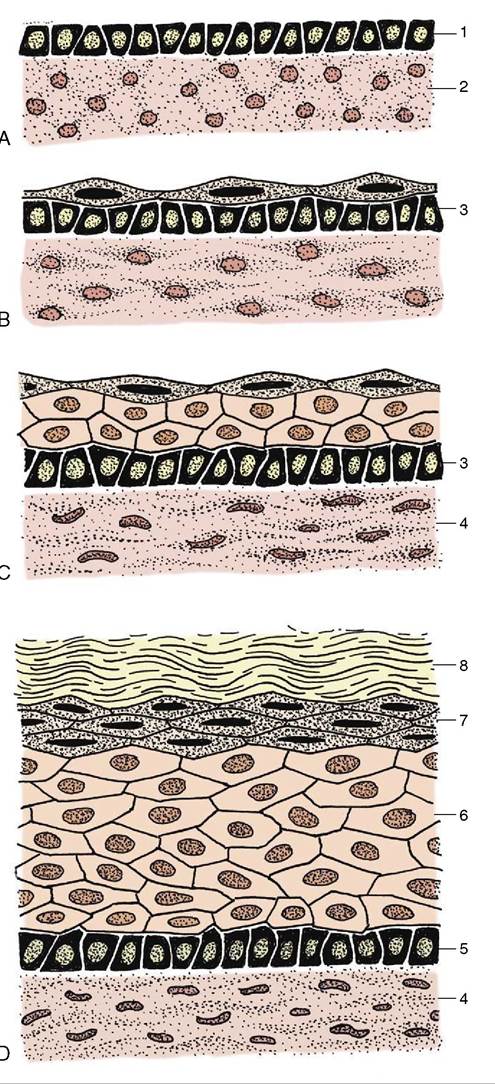
Figure 10-6 Development of skin, schematic. A, Skin of an early embryo. B, Differentiation of epidermis and dermis. C, Further differentiation of the epidermis. D, Complete differentiation of the epidermis and dermis. 1, Ectoderm; 2, mesoderm (mesenchyme); 3, primitive stratum basale; 4, dermis; 5, stratum basale; 6, stratum spinosum; 7, stratum granulosum; 8, stratum corneum.
Each hair grows from a tiny pit or follicle from which it protrudes above the surface of the skin.
The follicle develops from an ectodermal bud that grows into the underlying mesenchyme in the embryonic stage of life. In addition to forming the hair, the bud branches give rise to skin glands (Figure 10-7). The distal end of the bud forms a bulbous enlargement, which is then indented by a mesenchymal (dermal) papilla to form a primitive hair follicle. The epithelial cells lying against the papilla multiply, forming a hair matrix; the cells produced here keratinize and combine to form a primitive hair that grows through the center of the bud until it rises above the epidermis on the surface of the skin. Its passage takes it past the sebaceous glands that develop to the side of the follicle, and this arrangement allows the hair to obtain the oily coating so important for its health. While the ectoderm differentiates in this way, the mesoderm also condenses so that the tiny sheath around the embedded part of the hair acquires an outer mesodermal component.Figure 10-8 shows the essential features; other texts must be consulted for the histological details. It must suffice here to say that, in essence, a hair consists of a flexible column of closely consolidated and heavily keratinized, and hence dead, epithelial cells. Their arrangement permits the distinction of a medulla or core, a cortex, and an outer “scaly” cuticle. The proportions of the parts and the details of their arrangement vary and permit the microscopic determination of the origin of a hair sample. In general, hairs with a thick medulla are straight and rather brittle, whereas those in which the cortex predominates are stronger and more pliable.
The proximal end of the follicle is joined by a tiny arrector pili muscle passing from an attachment near the dermal papillae (Figure 10-7Zpronounced in wild species, but even domesticated animals protected from the more extreme climatic changes show
a recurrent pattern with peaks in the spring and fall. Shedding is obviously most noticeable in animals that are not regularly groomed to remove dead hair.
Information on these matters is not abundant, and most accounts rely heavily on casual observation. This is particularly so where companion animals are concerned, and veterinarians are frequently embarrassed by too- penetrating inquiry from owners. Although there seems to be much variation, most dogs molt most heavily in the spring and fall; the spring shedding is more pronounced and lasts about 5 weeks. Cats also molt most heavily in spring, but this is followed by a less substantial loss that continues through the summer and fall; it is not until winter that shedding ceases and the coat attains its prime condition. For the same reason, the pelts of furbearing species are harvested in winter, although the number of harvested pelts has been reduced as the trade in furs is regarded with increasing disfavor.The seasonal replacement begins with a slowing of the growth of existing hair; although this appears to be mainly conditioned by a rise in temperature, other factors, including nutrition and day length, play their parts. As growth slows (in the so-called catagen phase) the hair matrix and covering papilla both atrophy (Figure 10-9, B). No growth occurs in the ensuing (telogen) phase when the follicle, including the papilla, shortens, which causes a larger part of the hair to project above the skin in simulation of growth (Figure 10-9, D). When growth resumes, the follicle, with its matrix now reactivated, lengthens, and as it again extends away from the surface, it loses its grip on the old hair, which falls out. A replacement hair then forms in the active growth (anagen) phase that follows; this new hair gradually grows from the depth of the follicle until it emerges on the surface of the skin.
Wool hairs provide the soft undercoat. They are thin, wavy, and in most species, shorter and more numerous than the guard hairs by which they are concealed. The distinction between hair fiber types is not always clearcut, and intermediate forms exist to complicate description.
The sheep fleece presents particular problems as well as obvious interest. Wool is not, of course, confined to sheep among domestic animals. Cashmere and Angora goats, Angora rabbits, and alpacas all produce wools of distinctive quality that are utilized in the production of luxury yarns and textiles.In many species, including mature dogs and cats, several hairs share a single follicle opening (Figure 10-10, B-D). The central (primary) hair is longest and of the guard type, while the surrounding (secondary) hairs are shorter and softer; they provide the undercoat and may be designated wool hairs as they have little medulla.
The grouping of the hair follicles shows considerable interspecific and intraspecific variation. This may be revealed in products prepared from animal skin. The study of vellum of different periods has been used to trace the evolution of the fleece of modern breeds of sheep from the haircoat of their wild ancestors. Fragments of the Dead Sea Scrolls are among the materials utilized.
Tactile hairs are substantially thicker and generally protrude beyond the neighboring guard hairs. Most are found on the face, principally on the upper lip and about the eyes, although others are scattered (in species-variable fashion) on the lower lip, the chin, and elsewhere on the head. The cat, whose whiskers are particularly good examples (Figure 10-11), also possesses a cluster of similar hairs at the carpus. Tactile hair follicles reach deeply into the subcutis or even the superficial muscles. They are characterized by the presence of a venous sinus filled with blood and located between inner and outer layers of the dermal sheath (Figure 10-12). The nerve endings responsive to mechanical stimulation are also contained within the dermal sheath (Figure 10-12, A). The stimulus provided by disturbance of the hair is amplified by wave motion in the blood. The follicles of tactile hairs appear early in development, before those of the coat hairs, and their staged appearances provide useful criteria for aging embryos.
The skin of dogs and cats presents minute scattered tactile elevations (toruli tactiles) usually associated with special (tylotrich) guard hairs; the roots of these are surrounded by venous sinuses similar to, though smaller than, those of true tactile hairs. These elevations are also sensitive to touch (Figure 10-13).
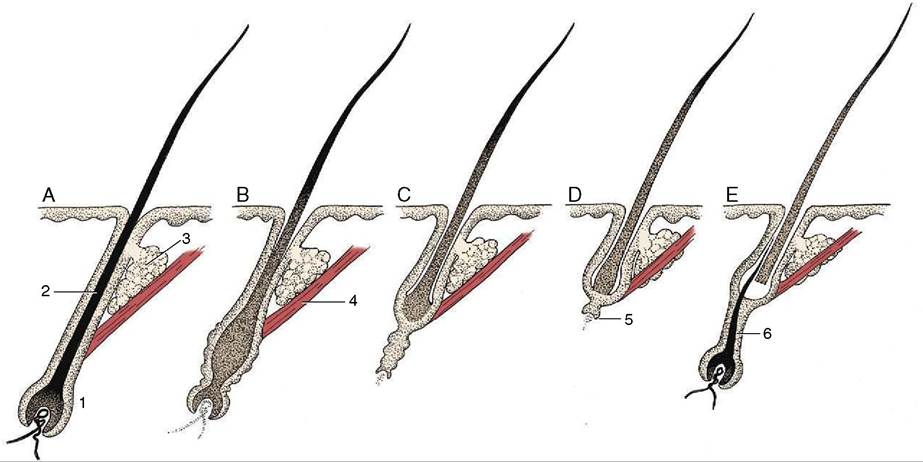
Figure 10-9 Phases of the hair cycle. A, Fully functional hair follicle; anagen phase. B, Follicle begins to atrophy; early catagen phase. C, Further atrophy of follicle; late catagen phase. D, Atrophied follicle. Hair is displaced distally and new hair matrix begins to form; telogen phase. E, New hair matrix established and new hair begins to grow; early anagen phase. 1, Hair follicle; 2, root of hair; 3, sebaceous gland; 4, arrector pili muscle; 5, new hair matrix; 6, new hair.
Many breeds of domestic animals, such as Holstein cattle and Dalmatian dogs, are immediately recognizable from the distinctive patterns of their coats. These patterns are created by the restricted distribution of various pigments: polymers of melanin ranging from black, through brown and red, to lighter shades that are present in granule form within cells of the epidermis, hair follicles, and hair. The pigments protect the skin from potentially harmful ultraviolet radiation and are unnecessary within those epidermal regions that are covered by a dense coat of hair. In most mammals, unlike humankind, skin pigmentation is therefore restricted to a few exposed parts that include the modified area associated with the external nose. It may be lacking here in white-coated individuals that obtain equivalent protection from a thickened stratum corneum.