Implantation and Placentation
Implantation is attachment of a blastula to the uterine epithelium and penetration of the epithelium by embryonic tissue. The degree of penetration by embryonic tissues varies among species.
in most domestic animals, the degree of penetration is much less than in rodents and primates, whose penetration extends into the connective tissue beneath the epithelium. implantation in domestic animals is considered to be noninvasive and primarily the result of formation of cell-to-cell junctions between embryonic tissues and the uterine epithelium. These junctions involve binding of membrane proteins in embryonic tissues to receptors on maternal epithelium. After fertilization, attachment occurs in the sow at about 11 days, in the ewe about 16 days, in the cow about 35 days, and in the mare about 55 days.Developing embryos migrate (i.e., move about) within the lumen of the uterus before implantation. in litter-bearing animals, this migration permits the spacing of embryos so that each has adequate room for development and ensures that each uterine horn contains some embryos. Contact, or paracrine communication, between embryos and the uterine epithelium is necessary for pregnancy recognition, and in litter-bearing animals, each uterine horn must contain embryos to permit this recognition. Embryos of non-litter-bearing species also migrate within the uterus before attachment. in the mare, migration back and forth between uterine horns before attachment is necessary to prevent luteolysis and loss of the developing embryo.
Placentation is development of the extraem- bryonic membranes, or placenta. The placenta is an arrangement of membranes with sites for exchanges between the maternal and fetal circulations so that nutrition from the dam can reach the fetus and waste products from the fetus can be transferred to the dam. in domestic animals, the terms fetal membranes and placenta are used interchangeably, although technically the fetal membranes are called the fetal placenta to distinguish them from maternal components of the placenta.
in some species a portion of the endometrium is also shed at parturition. This is the maternal placenta, or decidua. The fetal placenta includes the chorion, allantois, amnion, and vestigial yolk sac.The chorion, the outermost membrane, is in contact with the maternal uterine endometrium. The next layer (moving from outermost inward), the allantois, is a continuous layer that encloses a sac, the allantoic cavity (Fig. 28-2). The chorion and the outer layer of the allantois fuse to form the chorioallantois (Fig. 28-2). The amnion is the innermost membrane, closest to the fetus. It is a fluid-filled cavity that contains the fetus (Fig. 28-2). The amnion is fused with the inner layer of the allantois. The allantoic cavity, sometimes called the first water bag, is continuous with the cranial extremity of the bladder by way of the urachus, which passes through the umbilical cord. The fluid- filled amniotic cavity or sac is sometimes called the second water bag. The terms first and second water bag refer to fetal membranes at the time
Figure 28-2. Equine fetus in the placenta. The chorioallantois is the outer allantois plus the chorion. The chorion is associated intimately with the endometrium. The inner allantois is fused with the amnion. (Reprinted with permission of Wiley-Blackwell from Reece W.O. Physiology of Domestic Animals. 2nd ed. Baltimore: Williams & Wilkins, 1997.)
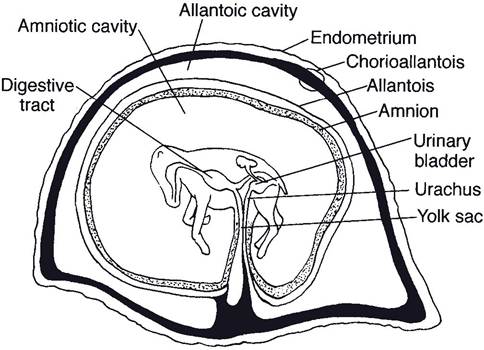
of parturition, when the allantoic sac is expelled first and the amniotic sac, second.
Branches of the umbilical arteries and veins run through the connective tissue between the allantois and chorion. These vessels are an important part of the fetal circulation. The umbilical arteries and their branches carry unoxygenated blood from the fetus to the placenta, and the umbilical vein carries oxygenated blood from the placenta to the fetus.
As a general principle, blood from the fetus never mixes with blood from the dam.
However, the two circulations are close enough at the junction of chorion and endometrium to permit oxygen and nutrients to pass from the maternal blood to the blood of the fetus and waste products to pass from the fetal blood into the bloodstream of the dam.The relationship between the fetal and maternal tissues at the histologic (microscopic) site of exchange is a basis for classification of mammalian placenta. The tissue of the maternal side is usually named first, then the fetal side. The placenta of most domestic mammals (sow, mare, ewe, and cow) is classified as epitheliochorial. In this type, the chorion of the fetus is in direct contact with the epithelium of the uterus of the dam (Fig. 28-3). (The placenta of ruminants may also be termed a synepithe- liochorial placenta because a number of fetal trophoblast cells fuse with endometrial cells to form binucleate cells.) A hemochorial type of
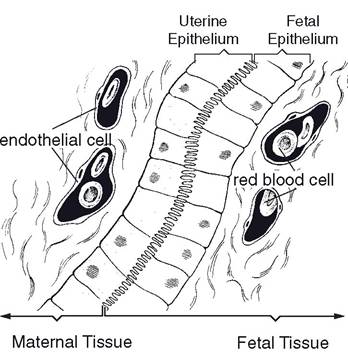
Figure 28-3. Epitheliochorial type of placenta.
placenta, in which fetal vessels and chorion are invaginated into pools of maternal blood, is found in humans and some rodents. An endotheliochorial type, in which the chorion is in direct contact with the endothelium of blood vessels of the dam, is found in carnivores. Mammalian placenta can also be classified at the gross anatomic level based on the distribution of the microscopic sites of exchange. in the mare and sow, extensions of the chorion (chorionic villi) project into crypts scattered over the entire endometrium, and this placental type is termed diffuse (Fig. 28-4). Ruminants have a cotyledonary type of placental attachment
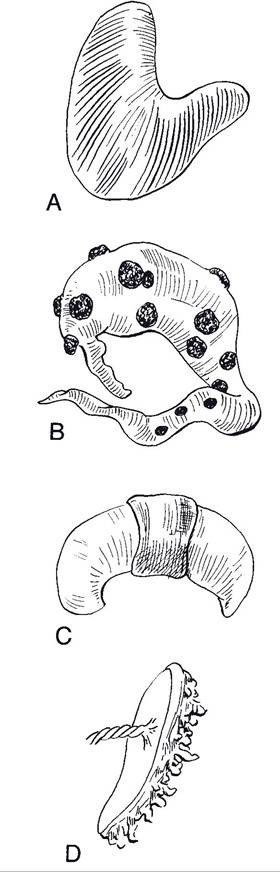
Figure 28-4. Placental types classified by distribution of sites of exchange. A) Diffuse placenta. B) Cotyledonary placenta. C) Zonary placenta. D) Discoid placenta. (Reprinted with permission of Wiley-Blackwell from Reece W.O.
Physiology of Domestic Animals. 2nd ed. Baltimore: Williams & Wilkins, 1997.)(Fig. 28-4) in which exchange takes place at structures termed placentomes. Placentomes are formed by the invagination of a specific region of fetal chorionic tissue, cotyledons, into a mushroomlike projection from the surface of the endometrium, caruncles (Fig. 28-5). These caruncles project out from the surface of the uterus approximately half an inch and vary in diameter from half an inch to more than 4 inches.
The size of caruncles increases as pregnancy progresses, and the caruncles are larger in the gravid (pregnant) horn than in the nongravid horn. The epithelial surface of the caruncle is covered with crypts into which villi of the fetal placenta project. The area between the caruncles is devoid of any attachment between the fetal placenta and the maternal uterus. The shape of the caruncles in the ewe is slightly different from those of the cow, having a rather large central depression that is the only portion of the caruncle to contain crypts for the attachment of the chorionic villi (Fig. 28-5).
Hemochorial placentas are usually attached to the uterus in a disk-shaped area only. Hence the term discoidal is used to describe their general area of attachment. The endotheliochorial placenta of carnivores is attached in a girdlelike band, so the attachment is known as zonary. The discoidal and zonary types of placental attachment are deciduate, since a portion of the maternal endometrium, or maternal placenta, is shed at the time of parturition. Most domestic animals have an indeciduate placenta, in which little or no maternal tissue is lost at parturition.