Junior management: Spinal cord and brainstem
Key points
■ Nerve fibres in the spinal cord enable local connections between sensory and motor nerves for reflex activity. They also provide links between the body and the brain for sensory and motor functions.
■ Both the grey and white matter can be divided functionally into distinct regions.
■ The dorsal funiculus carries ascending sensory tracts. The lateral funiculus carries ascending sensory tracts and descending motor tracts facilitating flexor muscle activity. The ventral funiculus carries mainly descending motor tracts facilitating extensor muscle activity. Each funiculus also transmits fibres connecting between spinal cord segments.
■ Some tracts remain ipsilateral whilst others decussate. Knowing whether a tract influences the ipsi- or contralateral side is clinically significant.
■ LMNs innervating the limbs are confined to the cervical and lumbar intumescences. The cranial part of the intumescence innervates cranial and proximal muscles of the limb. The caudal intumescence innervates the distal and caudal muscles of the limb.
■ UMN tracts in the spinal cord originate in the brain and mainly influence γ-LMN, via interneurons.
■ Nerve fibre diameter varies between tracts. Larger diameter fibres are more vulnerable to injury. Thus spinal cord compression sequentially results in proprioceptive dysfunction, paresis, autonomic dysfunction, loss of tactile sensation, paralysis and finally, loss of all nociception caudal to the lesion.
Overview of the spinal cord
The spinal cord has a variety of functions that can be summarised as the following:
1. Receiving and distributing information to the PNS;
2. Local integration (junior management) of sensory and motor functions for reflex activity, both within a limb and between limbs;
3. Relaying afferent/sensory information to the brain centres (senior management);
4.
Relaying efferent/motor information from senior motor management centres (UMN nuclei and motor cortex), via UMN tracts to connect with LMN;5. Forming connections between caudally directed tracts from the brain and cranially directed tracts, such that the former can regulate transmission of impulses in sensory systems. This is part of the mechanism by which nociceptive stimuli are modified.
The spinal cord has central grey matter and peripheral white matter. The grey and white matter can be divided functionally (Fig. 4.5). Each grey matter horn, or white matter funiculus, can be divided into distinct regions. There are ten different zones of the grey matter and more than a dozen specific tracts in the white matter. The details of those divisions are summarised in Fig. 4.5 and Table 4.2.
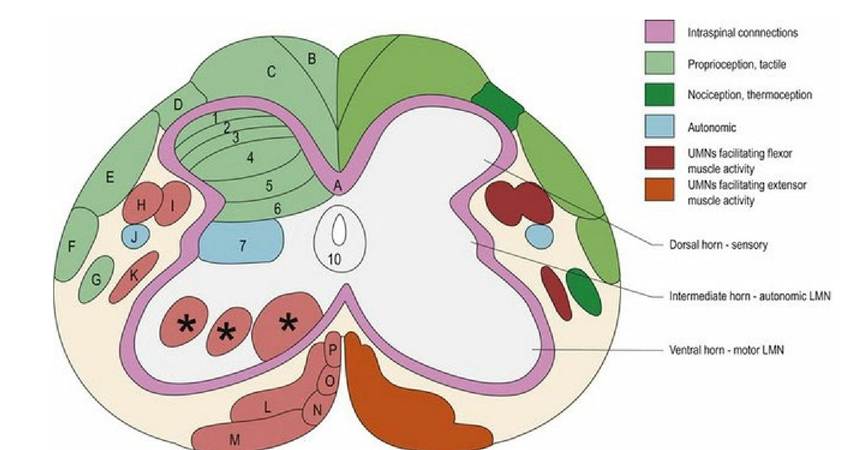
Fig. 4.5 Transverse section of spinal cord identifying grey matter zones and white matter tracts on one side and their general functions on the other. This is a hybrid diagram and the specific white matter tracts do not appear in all regions of the spinal cord. In the cervical region there are additional tracts, which are not depicted and there is no intermediate horn of grey matter (GM). Cranially and caudally directed tracts intermingle and are not as discrete as the diagram would imply. Grey matter is divided into ten zones (Rexed’s laminae) based on cellular architecture; this histological division has some correlation with function. The arrangement of all zones are variable throughout the cord, especially for zones 8 and 9; these are indicated by an asterisk (*). Details of tracts and laminae are given in Tables 4.2 and 4.3. Note that nociception is also conveyed in the dorsal and ventral funiculi as well as other parts of the lateral funiculi (see Chapter 6). Green colours = sensory, turquoise = autonomic, red colours = motor.
Table 4.2 The location and function of different regions of grey matter of the spinal cord; approximate correlations with the laminae are noted.
| Location | Grey matter column | Function |
| Top of dorsal horn (1,2,3) | Marginal nucleus and substantia gelatinosa | Nociception |
| DH deep to substantia gelatinosa (3,4) | Nucleus proprius | Interneurons and projection neurons |
| Base of DH, T1-L2 (5) | Visceral nuclei | Projection of visceral afferent stimuli to brain |
| Base of DH, T1-L4, medial aspect (5,6) | Nucleus thoracicus (also called nucleus of the dorsal spinocerebellar tract) | Projection of proprioceptive input from caudal half of body to cerebellum and nucleus Z |
| IH T1-L3 (7) | Intermediomedial and intermediolateral nuclei | Efferent, sympathetic presynaptic neurons |
| IH S1-S3 (7) | Sacral parasympathetic nuclei | Efferent, parasympathetic presynaptic neurons |
| Medial VH all segments excluding L7 and S1 (8,9) | Medial motor neurons | LMNs supplying truncal/axial muscles |
| Central VH C1-C6 (8,9) | LMNs of spinal accessory nerve (CN XI) and phrenic nucleus | Innervation of shoulder and neck muscles, and the diaphragm |
| Lateral aspect especially at the intumescences (9) | Lateral motor neurons | Innervation of the appendicular muscles |
| Surrounding the central canal (10) | Central intermediate substance | Nociception |
DH = dorsal horn, IH = intermediate horn, VH = ventral horn.
Spinal cord grey matter
The function of different areas of grey matter is outlined in Table 4.2.
The motor neurons of the lateral aspect of the ventral horn supplying the limbs are somatotopically arranged. Longitudinally, this arrangement means that cranial segments of the intumescence supply the cranial and proximal muscles of the limb and the caudal segments supply the caudal and distal muscles. This is clinically relevant. For example, a lesion in the cranial cervical intumescence (C6-C7) will damage the innervation to the shoulder muscles resulting in a shortened stride due to loss of shoulder movement. Damage to the caudal cervical intumescence (C8-T2) will compromise the radial, median and ulnar nerves resulting in failure to extend the elbow and bear weight on the limb and decreased movement of the carpus and digits.
Species differences include an additional nucleus (retrodorsal n.) located at C8-T1 and S1-S3 provides fine digital control in certain species, such as humans.
Spinal cord white matter
Within the CNS, nerve bundles form functional tracts that have common origins and destinations. Such tracts may also be called fasciculi (fasciculus - L = little bundle). The specific name of a tract, or fasciculus, usually indicates its origin and termination. For example, the vestibulospinal tract originates in the vestibular nuclei and ends in the spinal cord.
The white matter of the spinal cord is summarised in Table 4.3.
Table 4.3 The function of the white matter tracts indicated in Fig. 2.5
| Label | Tract name | Function: S = sensory, M = motor |
| A | Propriospinal (spinospinal) | Connections between spinal cord segments; used in reflex activity |
| B | Fasciculus gracilis | .... χ... S: Proprioception and tactile input from caudal of body. Post-synaptic tract intermingled conveying pinprick pain* |
| C | Fasciculus cuneatus | ..... /.... S: Proprioception and tactile input from cranial '' of body. Post-synaptic tract intermingled conveying pinprick pain*. Cuneocerebellar tract fibres at base of FC, conveying proprioception from cranial half of the body |
| D | Dorsolateral fasciculus | S: Nociception, thermal, multisynaptic |
| E | Dorsal spinocerebellar | S: Proprioception from body caudal to thoracic limb |
| F | Ventral spinocerebellar | S: Proprioception from caudal half of the body. The cranial spinocerebellar tract is found on the medial aspect and receives proprioceptive input from the cranial half of the body |
| G | Spinothalamic | S: Nociception, touch and temperature, multisynaptic These modalities are also conveyed by other tracts in lateral and ventral funiculi* |
| H | Rubrospinal | M: Flexor activity of body and limbs, limb protraction during locomotion |
| I | Lateral Corticospinal | M: Flexor and extensor activity especially of the distal limbs (voluntary, skilled movement leading to dexterity), also urinary bladder function |
| J | Lateral tectotegmentospinal | M: Sympathetic supply to head, e.g. for pupil dilation |
| K | Medullary (lateral) reticulospinal tract | M: Strongly inhibitory to extensor activity such as standing; it has a key role in flexor phase of gait. This tract arises from the medial medullary motor nucleus |
| L | bgcolor=white>Pontine (ventral)M: Facilitates extensor activity, e.g. standing and antigravity muscles; key role in extensor phase of gait | |
| M | Lateral vestibulospinal | M: Vestibulospinal: Ipsilateral extension, inhibition of ipsilateral flexion and contralateral extension; key role in extensor phase of gait |
| Spinovestibular | S: Spinovestibular: proprioception of head-neck junction to vestibular nuclei | |
| N | Tectospinal | M: Contraction of neck muscles for reflex turning of head in response to auditory and visual stimuli |
| O | Ventral corticospinal | M: Facilitation of voluntary movement in the neck and trunk |
| P | Medial vestibulospinal and medial longitudinal fasciculus | M: Ipsilateral extension of neck muscles and inhibition of contralateral extension |
* See Chapter 6 and Table 5.1 for motor tracts
There is a general pattern to the functions conveyed by spinal cord white matter. Each funiculus has the propriospinal tract making intersegmental connections between cranial and caudal spinal cord segments. The dorsal funiculus carries ascending sensory tracts; the lateral funiculus carries both ascending sensory tracts and descending motor tracts facilitating flexors and the ventral funiculus carries mainly descending motor tracts facilitating extensors. However, it should be noted that there are some exceptions to this general pattern. For example, the following cranially directed tracts are found in the ventral funiculus.
1. Spinoreticular tracts - sensory and nociceptive input to the reticular formation and then to the thalamus and cerebrum, for alerting and motivational purposes.
2. Spinovestibular tracts - proprioceptive input from the cervical region to the caudal vestibular nuclei (head proprioception).
3. Spinomesencephalic tract to midbrain and the thalamus. This tract may be involved in the activation of a caudally directed analgesia system.
4. Spino-olivary fibres - convey sensory feedback to the olivary nucleus and hence cerebellum. Spino-olivary fibres are found in the spinoreticular tract (VF) the ventral spinocerebellar tract (LF) and the fasciculus cuneatus (DF).
There are also other caudally directed tracts in the lateral funiculus, such as fibres from the locus ceruleus, which is a collection of adrenergic neurons in the pons, and the raphe nuclei of the rostral medulla oblongata. Both tracts are involved in modulating the response to noxious stimulation, the first by releasing noradrenaline and the second by releasing serotonin.
Note that tracts and pathways are not the same. A tract, or fasciculus, is a specific bundle of axons in the CNS extending from dendrites to synapse. Tracts are usually named for their origin and termination, e.g. the vestibulospinal tract. A pathway is a route through the nervous system that may involve several tracts, synapsing successively with each other. For example, the proprioceptive pathway from the thoracic limbs is as follows: afferent impulses from the PNS travel cranially in the spinal cord via the fasciculus cuneatus, synapse in the cuneate nucleus of the medulla oblongata, travel via the medial lemniscus to the thalamus, synapse and then go via thalamo-cortical fibres to the somatosensory cortex.
Many tracts decussate (e.g. corticospinal, rubrospinal tracts) meaning they cross the midline in the CNS, thus the origin and termination of the tract are on opposite sides.
Spinal cord functions: Links with the PNS
During embryonic development the neural tube has a relatively uniform diameter along its length. However, if a neuron fails to connect to a target, the neuron degenerates. There are more peripheral targets in limb regions, hence in the postnatal animal the spinal cord diameter is larger in regions supplying the limbs as it houses more nerve cell bodies and nerve fibres. These areas of increased diameter are called the cervical and lumbar intumescences (tumere - L = to swell up). In a transverse section of the spinal cord at an intumescence, the dorsal and ventral horns are enlarged (more nerve cell bodies) and the nerve roots are more prominent (Fig 4.6A, B).
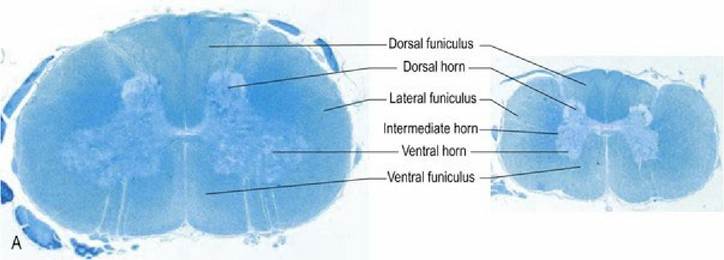
Fig. 4.6A Dog spinal cord, transverse sections, at the levels of the cervical intumescence (left) and cranial lumbar spinal cord (right). Note the increased size of the dorsal and ventral horns in the cervical intumescence compared with the lumbar cord and the presence of an obvious intermediate horn supplying autonomic efferent fibres to the viscera, in the latter. Spinal cord sizes are in proportion with each other; this illustrates that the cord diameter is larger at the intumescence. Spinal cord sections stained with luxol fast blue and cresyl violet.
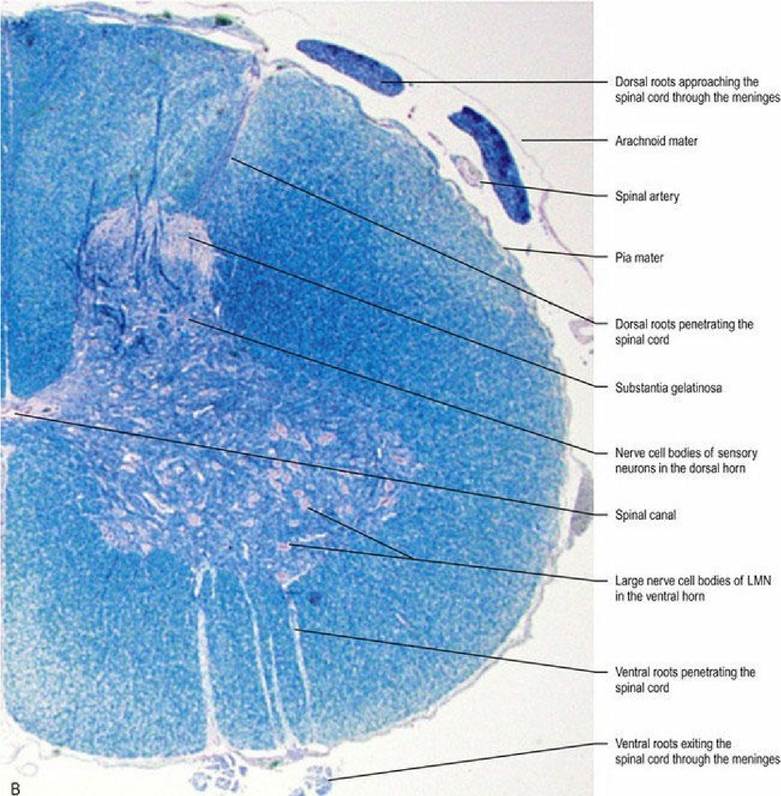
Fig. 4.6B Higher magnification of the C7 cord segment.
Longitudinally, the different regions of the intumescences innervate different aspects of the limbs, via the brachial or lumbsosacral plexi (see Fig. 1.2). The pelvic plexus innervates pelvic viscera and the perineum. Spinal nerves attached to the cranial part of the intumescence will mingle in the cranial part of the associated plexus and then innervate the proximal and cranial muscles of the limb. Spinal nerves attached to the caudal part of the intumescence will mingle in the caudal part of the associated plexus, and then innervate the distal and caudal muscles of the limb. This leads to a general rule that the cranial intumescence and plexus innervate cranial and proximal muscles of the limb, whilst the caudal intumescence and plexus innervate the caudal and distal muscles of the limb. Knowing which spinal cord segments contribute to the intumescence allows the clinician to localise the lesion based on observed signs. For example, observing that the supraspinatus muscle is atrophied and the animal cannot extend the shoulder would indicate a lesion in the cranial part of the brachial plexus or the cervical intumescence (Fig. 4.7).
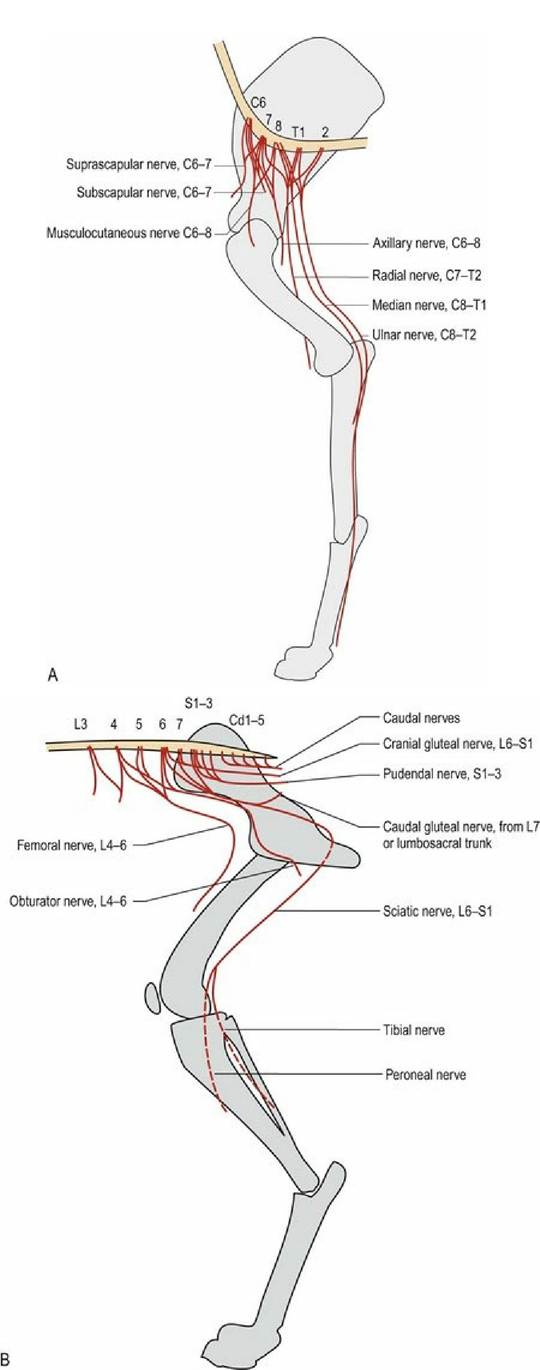
Fig. 4.7 Innervation of the thoracic and pelvic limbs from the intumescences via the plexi, viewed from the medial aspects. The cranial intumescence and plexus innervates the proximal, cranial muscles of the limb.
The caudal intumescence and plexus innervates the distal and caudal muscles of the limb.
Clinically, the spinal cord can be divided into functional areas, based around which cord segments supply the limbs, into the C1-C5, C6-T2, T3-L3, L4-S3 and Cd1-5 regions for the dog (see Fig. 1.2). Knowing these divisions is key to interpreting the clinical neurological examination of the spinal cord (see Chapter 13).
A lesion in any region of the cord can affect both the grey matter and white matter. Damage to grey matter causes loss of neurons and their axons will degenerate. Clinically, this is particularly important if the lesion affects the nerve cell bodies of the LMNs in an intumescence. Their LMN fibres connecting to the NMJ in the limb muscles will degenerate and the muscles will atrophy. Damage to other nerve cell bodies in the grey matter, or their axons in the white matter, will lead to loss of axons distal to the lesion. If sensory somata, or ascending nerve fibres, are damaged, axons cranial to the lesion will be lost. Conversely damage to neurons whose axons connect to caudal segments (e.g. for reflex function) or damage to UMN fibres, will cause loss of axons caudal to the lesion. These patterns of damage can be recognised histopathologically. Damage to white matter can cause loss of sensation or motor function, resulting in anaesthesia and paresis/paralysis caudal to the site of the lesion.
Localising lesions in the spinal cord revolves around evaluating spinal Reflexes, muscle Atrophy and Tone - or the ‘Neuro RAT’ (see Fig. 5.6 and Table 5.2). Incoming sensory fibres from the thoracic and pelvic limbs enter the cervical or lumbar cord segments, respectively. Those fibres can synapse with the LMNs supplying that limb, which then connect via the NMJ to striated muscles in that limb. Thus, as long as those spinal cord segments are functioning normally, reflex arcs will be intact and the limb muscles will have tone. If the spinal cord segments supplying the limbs are damaged, this can cause loss of reflex function, loss of tone and marked wasting of the muscles (neurogenic atrophy) due to loss of the LMN (‘LMN lesion’) and degeneration of the neuromuscular junction. In contrast, damage to the segments cranial to the intumescence can result in deficits in the body and limbs caudal to the lesion. The animal may have sensory deficits and, due to damage to UMN (‘UMN lesion’), paresis/paralysis. However, the reflexes and limb muscle tone will still be intact in the limbs caudal to the lesion and any atrophy will be mild and due to disuse of the muscle. Note that reflexes and tone may actually be increased with such lesions due to damage to UMNs that are inhibitory to LMN.
Spinal cord functions: Links within the spinal cord
Sensory input from the periphery enters the cord via the dorsal root and may synapse on interneurons in the dorsal horn or pass through the dorsal horn into the white matter or ventral horn without synapsing. Output from the dorsal horn may:
I Link to LMNs in the ipsilateral ventral horn of the same spinal cord segment for reflex function;
II Decussate in the white commissure to the contralateral ventral horn of the same spinal cord segment for reflex function;
III Enter the white matter to travel cranially or caudally to another spinal cord segment, ipsilateral or contralateral to the original side, and synapse with LMNs for reflex function;
IV Enter the white matter to travel cranially to the brain.
Interneurons are involved in all of these connections, excluding the monosynaptic relay found in some myotatic/stretch reflexes (Fig. 4.8). Axons that connect between spinal cord segments form the propriospinal tract; this is also known as the spino-spinal tract as it starts and finishes within the spinal cord.
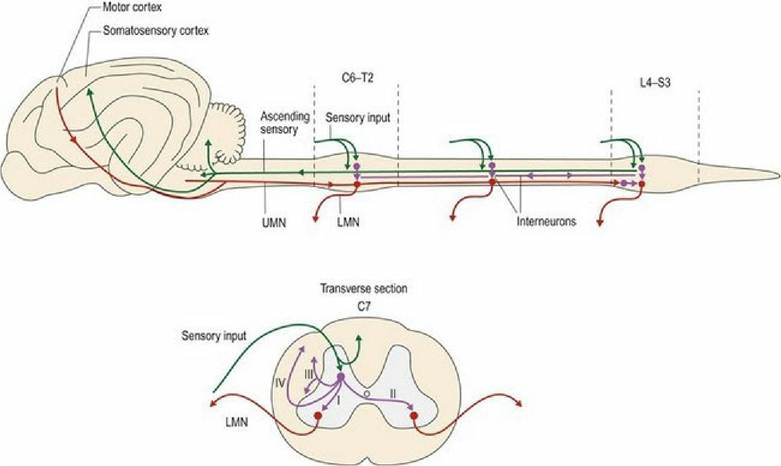
Fig. 4.8 Links within the spinal cord.
By virtue of links described in I-III above, input from one limb can result in reflex activity within that limb, utilising just one, or many, of the spinal cord segments involved in that intumescence. It can also affect the contralateral limb and the limbs of the other girdle. For example, stepping on a sharp object with the left pelvic limb will cause reflex withdrawal of that limb and simultaneous extension of the contralateral pelvic limb and potentially, the thoracic limbs too to accommodate the change in weight distribution.
Locomotion involves synchronised flexion and extension of the limbs, much of which is coordinated through wiring that forms reflexes (see Chapter 9), thus, it also utilises links described in 1-3. Events in each limb are linked temporally and spatially, to events in the other limbs. The basic circuitry for coordinated, rhythmical flexion and extension of the limbs is located in the spinal cord. But the spinal cord circuitry alone is insufficient for gait; input from brain centres is also required.
Spinal cord functions: Links to the brain
Most incoming sensory fibres from the PNS synapse in the dorsal horn. Fibres make local connections for reflex function or enter the white matter to travel cranially to the brain in specific, defined tracts within the dorsal or lateral funiculi. Such cranially directed fibres may or may not synapse in the dorsal horn first. Once they reach the brain they may: (a) terminate by synapsing in the brainstem on specific nuclei; (b) travel to the cerebellum without synapsing en route; (c) synapse in relay nuclei in the brainstem, from which they continue to the somatosensory cortex of the forebrain.
The UMN tracts in the spinal cord link the UMN motor centres of the brain with the LMNs of the spinal cord. The majority of UMN nuclei important for quadrupedal movement are located in the brainstem. The UMN input from the motor cortex of the forebrain directs voluntary, learned movements. The UMN systems initiate, modulate and terminate LMN activity and reflex function for posture and locomotion, voluntary movement and visceral function.
Most UMN axons in the spinal cord synapse on interneurons, which then synapse onto the LMN. There are some exceptions to this rule, depending on the species. The corticospinal tract and vestibulospinal tracts may synapse directly onto the LMN. Whether a UMN synapses directly or indirectly on LMNs is relevant clinically as indirect synapses onto diverse interneurons allow for divergence and convergence of information (Fig. 4.9). If there are several synapses, loss of a component of the pathway may be compensated for by rerouting the information through other neurons. The nervous system is relatively plastic and co-opting of alternative routes may assist in recovery after spinal cord injury. Conversely, if the synapse of the UMN is directly onto the LMN, then there are no alternative routes, using interneurons, by which information can get to the LMN. Humans have a much greater dependence on direct connections; this is one reason why a cerebrovascular accident (‘stroke’) affecting blood supply to the motor cortex or its efferent pathways, can cause significant motor deficits.
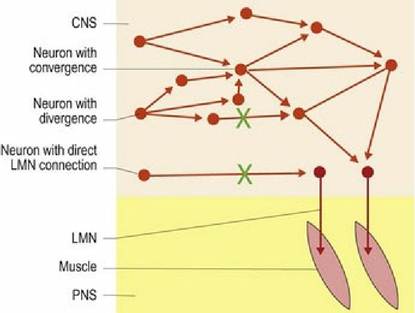
Fig. 4.9 Upper motor neurons connect directly or via interneurons to LMNs. Lesions as depicted by the green ‘X’ will result in complete loss of LMN control if UMNs make a direct link from the motor centre in
the brain. However, those that connect via interneurons may be able to reroute information and thereby still control the LMN.
Many tracts decussate, such that their axons cross the midline, either near their origin, e.g. rubrospinal tract, or near their termination, e.g. ventral corticospinal tract, thus tract input and termination are on the opposite sides of the body. Decussation of the nervous system probably evolved from the development of the coiling reflex in primitive chordates. A noxious stimulus on one side results in contraction of contralateral muscles, thus moving the animal away from the stimulus. A few tracts may cross the midline more than once, e.g. ventral spinocerebellar tract. Other tracts mainly influence the ipsilateral side, e.g. vestibulospinal tract. Sensory tracts travelling to the cerebellum ultimately terminate in the ipsilateral cerebellum. Sensory tracts travelling to the cerebrum, remain ipsilateral in the spinal cord, but after synapsing in the brainstem, their rostral continuation crosses to the opposite side. The exceptions to this rule are tracts like the spinothalamic tract that conveys nociception in quadrupeds. The spinothalamic tract travels cranially on both sides of the spinal cord in lateral funiculi, and in and out of the grey matter synapsing multiple times.
Decussation is clinically relevant as indicated by the example of an animal with a lesion that affects the pathway conveying touch and proprioception. Consider the cat in Fig. 4.10 that is bearing weight on the dorsal aspect of the right forepaw indicating abnormal paw position sense. The proprioceptive pathway originates in tactile and joint receptors of the forepaw, travels via the nerves (e.g. the radial nerve) into the dorsal horn of the C7-T1 spinal cord segments, into the dorsal funiculus (fasciculus cuneatus) cranially to the medulla oblongata where it synapses (nucleus cuneatus). From there the pathway decussates (deep arcuate fibres) and continues rostrally (medial lemniscus), synapsing in the thalamus and hence to the somatosensory cortex on the left side of the brain. This cat’s signs could be due to a lesion anywhere along that pathway from origin to termination. If the lesion were in the spinal cord, it would have to compromise the right dorsal funiculus, but if it were in the forebrain or rostral brainstem, it would have to be on the left side. Localising the lesion depends on the results of the rest of the neurological examination. For example, if the lesion was in the spinal cord, there may be signs of ipsilateral limb paresis, while if it was in the forebrain there may be signs of contralateral visual deficit (see Fig. 13.1).
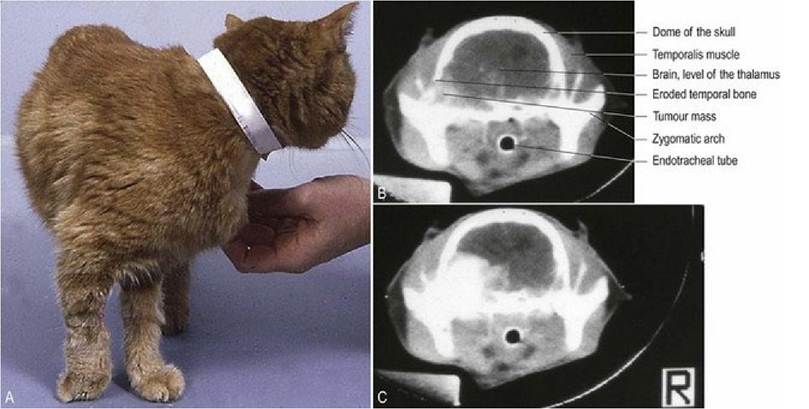
Fig. 4.10 (A) Cat with abnormal paw position due to a lesion affecting termination of the proprioceptive tract; specifically the cat had a tumour in the left forebrain. The cat was not paretic, but it did have a visual deficit in the right eye. The majority of the visual pathway from the right eye decussates in the optic chiasm ventral to the diencephalon (forebrain) to the left visual cortex. Hence clinical signs in the right side of the body were due to a left forebrain lesion. This contralateral location of the lesion relative to the clinical signs reflects decussation of the involved sensory pathways. Precontrast (B) and post-contrast enhanced (C) CT scans depicting a large extradural mass on the left side of the cranial vault.
The function of spinal cord tracts, and their vulnerability to compressive lesions, is related to their microanatomy; this vulnerability explains the sequence of neurological signs observed in compressive spinal cord lesions. Factors determining vulnerability are axonal diameter, thickness of the myelin sheath and fibre location in the cord. The conduction velocity of a nerve fibre is proportional to both the diameter of the fibre and the degree of myelination. Thus large-diameter, heavily myelinated fibres are fast conducting (e.g. myelinated, 20 μm diameter fibres conduct at velocity of up to 120 m/s), while small, lightly or nonmyelinated fibres are slow conducting (e.g. 1 μm diameter conducting at 1 m/s). Examples of such fibres are proprioceptive and nociceptive fibres, respectively.
In compressive lesions of the spinal cord, the first fibres to be compromised are the proprioceptive fibres, as they are large diameter and heavily myelinated. The last fibres to be compromised are those conveying nociception. That it takes a severe, extensive lesion to cause nociceptive dysfunction reflects the fact that nociceptive fibres are small diameter and lightly myelinated and, also, that there are several tracts that convey nociception; these tracts are located in different areas of the spinal cord (see Chapter 6). Other sensory fibres and motor fibres have axonal diameters and myelin sheath thickness that is between these two extremes.
For compressive spinal cord lesions of increasing severity, the signs will begin with spinal hyperpathia/sensitivity (due to meningeal irritation), proprioceptive dysfunction, then paresis, loss of superficial (pinprick) pain, paralysis, and finally loss of all nociception even from an intense crushing stimulus such as applied across a digit by a pair of large haemostats. Loss of urinary and faecal continence occurs when the spinal cord damage is severe enough to cause marked paresis or paralysis.