Membrane Fusion Allows for a Combination of Compartmentalization and Transport of Material
Impermeable molecules can be transported across the cell membrane by means other than membrane proteins. This method involves using membrane itself as a carrier compartment. The lipid bilayer of biological membranes has a structure similar to soap bubbles.
As with soap bubbles, small vesicles of biomembrane (essentially “membrane bubbles”) can fuse to form larger membrane surfaces. A large membrane surface can also pinch off (requiring fusion of two membrane surfaces) into small vesicles. When these processes occur at the plasma membrane, they are called exocytosis and endocytosis, respectively (Figure 1-11). VXTien these processes occur at internal membrane sites, the process is referred to as membrane fusion, whatever the direction. Membrane fusion underlies much of membrane vesicle traffic around the cell. This traffic creates intracellular vesicles, renews plasma membrane by adding newly synthesized membrane, and transports material within the cell and across the plasma membrane. Because the transport is compartmentalized within a membrane bubble, the transported material can be targeted specifically to one or another region of the cell. Also, changes to the “cargo” can occur within a particular membrane compartment, as seen in cholesterol transport.Exocytosis and endocytosis are crucial in the transport of cholesterol (Figure 1-12). Cholesterol is an essential lipid component of many animal biomembranes; the plasma membrane lipids of animals are about 15% cholesterol and 60% phospholipids. Cholesterol is also the starting material for the synthesis of the entire group of hormones called
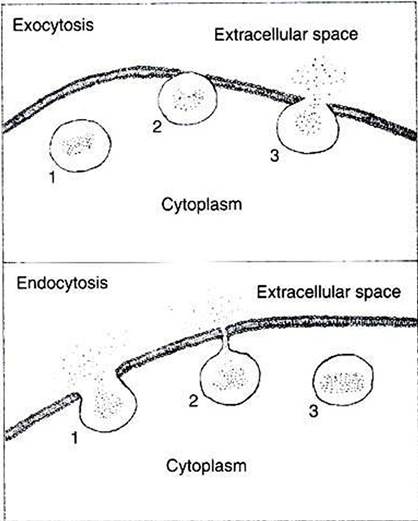
FIGURE 1-11 Two membrane fusion processes: exocytosis and endocytosis. Top, In exocytosis, a membrane-bound vesicle from the cytoplasm (1) makes contact and fuses with the plasma membrane {2).
As the vesicle membrane becomes continuous with the plasma membrane, the contents of the vesicle are released to the extracellular space (3). Bottom, In endocytosis, some material from the extracellular space is surrounded by plasma membrane (1)l which continues to invaginate until the edges are able to fuse (2)t thus pinching off a vesicle from the plasma membrane (3}. Membrane fusion can occur between any two compartments within cells separated by lipid bilayer membrane, not only between the cytoplasm and extracellular space, as shown here.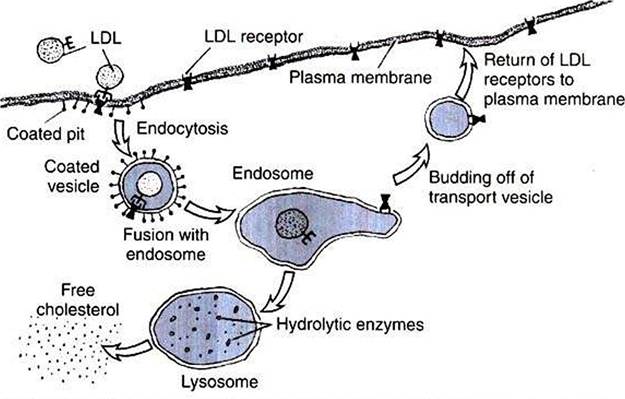
FIGURE 1-12 Processes of membrane fusion involved in cholesterol uptake by cells. Starting at the left, a low-density lipoprotein (LDL^containing cholesterol binds to an LDL receptor protein of the plasma membrane and undergoes endocytosis, forming an endosome.The receptor is detached from its LDL ligand in the endosome.The LDL portion of the endosome fuses with a lysosome to digest the LDL and produce free cholesterol, while the receptor-containing portion of the endosome pinches off a vesicle to return to the plasma membrane, thus recycling the receptor. (Redrawn from Alberts B, Bray D, Johnson A, et al: Molecular biology of the cell, NewYork, 1983, Garland.)
steroids (see Chapter 33). Cholesterol can be synthesized by animals and is also absorbed by meat-eating animals from their diet. Because cholesterol is soluble in oil, it passes from food through the plasma membrane without protein mediation into the cells of the gut lining. However, transport of dietary cholesterol through the circulatory system requires that cholesterol molecules form a complex with a protein molecule to create low-density lipoproteins (LDLs). To take up cholesterol from the circulation, cells bind the LDLs to intrinsic membrane proteins that act as LDL receptors, as shown in Figure 1-12. The receptor/LDL complex then diffuses in the plane of the membrane into specific regions to form coated pits.
The coated pit is taken into the cytoplasm by endocytosis. In addition to the transport function, receptor- mediated endocytosis functions also to concentrate extracellular material before internalization. The coated pit is not taken into the cell until it has collected the LDLs from a much larger volume of ECF than the cell could “drink." The membrane vesicles formed by this endocytosis fuse subsequently to become an endosome. The endosome compartment becomes acidic, which causes dissociation of the LDL and the receptor. Through unknown means, the endosome is then able to further separate and compartmentalize the receptor from the LDL. Membrane vesicles containing the now-vacated LDL receptors return to the plasma membrane and fuse by exo- cytosis. The LDL receptor is recycled to the plasma membrane to pick up more LDL. Experimental evidence suggests that a single LDL receptor molecule can cycle between the plasma membrane and endosomal vesicles more than IOO times before losing its activity. Meanwhile, the LDL moiety is segregated to another endosomal vesicle, which fuses with the lysosome. The lysosome contains hydrolytic enzymes, thus allowing the internalized LDL to be digested. The cholesterol is now available to the cell for steroid synthesis or incorporation into membrane.Other molecules are also recycled by endocytosis. As with the LDL receptor, for example, many signal receptors, discussed in the next section, are endocytosed back into the cell that released them, saving the cell the effort of synthesizing new receptors. Not all endocytosed molecules are recycled. Manv are broken down after their endosome fuses with a lysosome. Indeed, as described later, this is one method of regulating receptor number on the plasma membrane.