Membrane Potentials and Excitable Cells
Resting Membrane Potential
There is a relatively small difference in the amounts of charged ions locally on the opposite sides of the outer cell membrane of all animal cells. In most conditions (i.e., resting conditions), the outside of the cell membrane has a small excess of positive ions (cations), and the inside of the cell membrane has a small excess of negative ions (anions).
The excess negative and positive charges tend to attract each other, so they line up on each side of the membrane, creating an electrical potential across the membrane (Fig. 2-11). The measurable voltage difference is the membrane potential. The size of the membrane potential varies among types of cells from -10 mV (millivolts) to -100 mV. In many nerve and muscle cells, it is about -85 mV. This means that the inside of the membrane is 85 mV more negative than the outside.The concentrations of various cations and anions throughout the intracellular and extracellular fluids are maintained relatively constant in normal, healthy animals. As a result, two features of the membrane primarily determine the magnitude of the membrane potential. These are the transport mechanisms available to move cations and anions across the membrane and permeability of the membrane to the different ions.
Recall that all cell membranes contain the Na-K pump or Na-K-ATPase system (Fig. 2
14). The net effect of the Na-K-ATPase system is constant movement of Na+ out of the cell and K+ into the cell. The system actually moves out three Na+ ions for every two K+ ions that move in. This difference contributes to the net negative charge found on the inside of the membrane.
In resting conditions, cell membranes are relatively impermeable to Na+ (and proteins, which tend to be anionic) but are quite permeable to K+.
Even though some Na+ tends to leak back into the cell down the concentration gradient, the relatively low membrane permeability to Na+ and the continuous operation of the Na-K pump maintain the intracellular concentration of Na+ (10 mEq/L) less than that in the extracellular fluid (140 mEq/L).In contrast, the intracellular potassium concentration (140 mEq/L) is much greater than its extracellular concentration (5 mEq/L). Because the cell membrane is quite permeable to K+, it can freely diffuse out of the cell down the concentration gradient. This exit of a positively charged cation is a major contributor to the relative excess of negatively charged ions on the inside of the membrane. The importance of this exit of K+ is illustrated by the effects of changes in the concentration of extracellular K+ on the electrical activity of the heart. Abnormal increases in extracellular K+ concentration, such as may occur with kidney disease, are often associated with abnormal electrical activity of the heart (cardiac arrhythmias), and these can threaten life.
In the typical cell in resting conditions, sodium and potassium are the major determinants of the membrane potential. However, in some cell types and in certain conditions, the membrane permeability to other ions, hence passage of these ions across the membrane, may be a significant contributor to the membrane potential. For example, if the membrane permeability to Cl- increased, Cl- would diffuse into the cell down the concentration gradient, and the inside of the cell would become more negative.
Excitable Cells and Action Potentials
Nerve and muscle cells are excitable cells. In response to the proper stimulus, their cell membrane potential can undergo a rapid but short-lived reversal in electrical potential, so that the inside is positive to the outside. This event is known as an action potential (Fig.
215). The reversal of the resting membrane potential is described as depolarization of the membrane, because during this period the membrane potential is closer to zero. stimuli that elicit action potentials can be physical, chemical, or electrical. When a stimulus is strong enough to cause an action potential, it is a threshold stimulus.
The changes in membrane potential during an action potential are due to rapid changes in membrane permeability to different ions and movement of those ions across the membrane. The rapid changes in permeability are due to the opening and/or closing of membrane channels (i.e., channels formed by transmembrane proteins). The channel itself may respond to physical, chemical, or electrical stimuli, and the response of the channel is the link between threshold stimuli and action potentials. A
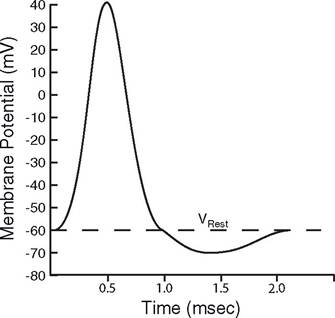
Figure 2-15. Nerve action potential.
threshold stimulus is one that causes enough change in channel status to bring on a rapid reversal of the membrane potential.
When an action potential occurs at a single site on an excitable cell, the membrane potential of adjacent areas along the membrane of the same cell also changes. The change in potential in the adjacent area is due to the movement of charge (ions) between the two areas (Fig. 2-16). If the membrane in the adjacent area has membrane channels sensitive to changes in voltage (voltage-gated channels) and if the change in membrane potential in the adjacent area reaches a threshold voltage, another action potential in the adjacent area is elicited. This can occur again and again, so that a series of action potentials moves along the surface of a cell (Fig. 2
16). This movement of action potentials is propagation of the action potential. This type of propagation is typical of all excitable cells (muscle and nerve cells).
Disrupting the activity of membrane channels can prevent action potentials and their propagation. For example, propagation of action potentials in peripheral nerves depends on the rapid opening of Na+ channels. Local anesthetics, such as lidocaine,
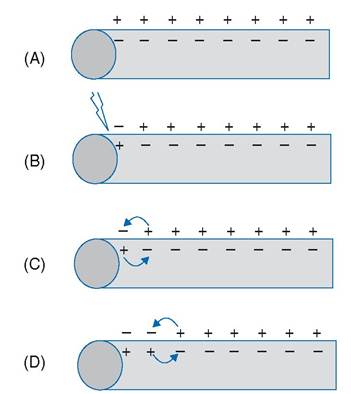
Figure 2-16. Propagation of action potential. A) Resting membrane potential. B) Initial threshold stimulus. C and D) Propagation.
block these sodium channels and prevent action potential propagation when applied along a peripheral nerve.