Membrane Proteins That Serve the Triple Functions of Selective Transport, Catalysis, and Coupling Can Pump Ions and Molecules to Regions of Higher Free Energy
Vant Hoff’s law and Starling’s hypothesis deal with passive transport (i.e., movement of material in the direction of lower electrochemical potential). However, the cell moves many ions/molecules against their electrochemical potential.
That is, this selective transport requires the expenditure of energy by the cell. Transport in a direction requiring an expenditure of energy (i.e., input of work) is called active transport. Active transport depends on intrinsic membrane proteins that use specific binding and allostery to achieve the dual functions of selective transport and reaction coupling. Many (but not all) active transport proteins obtain the energy for transport from ATP hydrolysis; these proteins must function also as enzymes (ATPases).An important example of active transport is the Na∖K' pump (also known as Na',K -ATPase). This intrinsic membrane protein consists of four polypeptide chains (2 a + 2 β) and has a mass of approximately 300,000 daltons. This molecule catalyzes the hydrolysis of Al P and couples the hydrolysis energy to the movement of Nat out of the cell and K into the cell. This ion pump creates and maintains a considerable concentration gradient across the cell membrane for both ions (see Table 1-1).
Figure 1-8 shows our current understanding of this protein’s structure and outlines the cycle of binding and conformational changes underlying its transport function. T'he Na∖K,-ATPase pumps three Na' ions out o/'the cell and two K* ions into the cell for each ATP molecule hydrolyzed. These directions of ion pumping cause a high Na' concentration outside the cell and a low concentration inside, whereas K* concentration is high inside and low outside the cell. The different directions of pumping for the two ions depend on differing binding specificity of the pump protein in the two
Table 1-1
Concentrations of Various Substances in the Intracellular, Extracellular, and Plasma Fluids
| Concentration (mmol∕L) | |||
| Intracellular | Extracellular | Blood plasma | |
| Na’ | 15 | 140 | 142 |
| K* | 150 | 5 | 4 |
| CA2* | 0.0001 | 1 | 2.5 |
| Mg2* | 12 | 1.5 | 1.5 |
| Cl | 10 | 110 | 103 |
| HCO3- | 10 | 30 | 27 |
| Phosphate | 40 | 2 | 1 |
| Glucose | 1 | 5.6 | 5.6 |
| Protein | 4.0 | 0.2 | 2.5 |
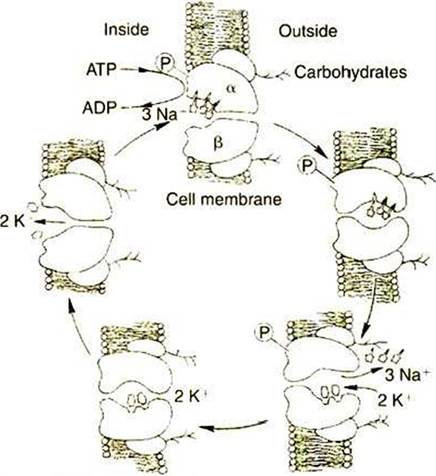
FIGURE 1-8 Hypothetical transport cycle for
Na*,IC-ATPase.
Changes in the conformation of this transport protein driven by ATP hydrolysis and ion-binding events cause three Na* ions to be moved out of the cell against a concentration gradient and two K* ions to be moved into the cell, also against a concentration gradient, for each ATP hydrolyzed. (Redrawn from a diagram by Dr. Seth Hootman.)conformational states. The ability of the protein to couple this transport to the enzymatic breakdown of ATP allows the transport to occur against the concentration gradients, from lower to higher electrochemical potentials for both ions. In the particular case of the Na,K pump, the number of transported electrical charges is asymmetrical; three positive charges leave for each two positive charges that enter. This asymmetry of electrical charge transport means that the Na+,K+ pump is electrogenic, making a minor contribution to the electrical potential (voltage) across cell membranes, as discussed later.
Many different intrinsic membrane proteins actively transport a wide variety of ions and molecules against the transported molecules’ electrochemical gradient. Many, such as the Na+,K* pump, couple the energy-requiring “uphill” transport with the “downhill” hydrolysis of ATR However, any potential source of free energy can be coupled to the energyrequiring transport. Indeed, the gradient of Na* set up by the Na*,K+ pump is itself used frequently as a source of energy.
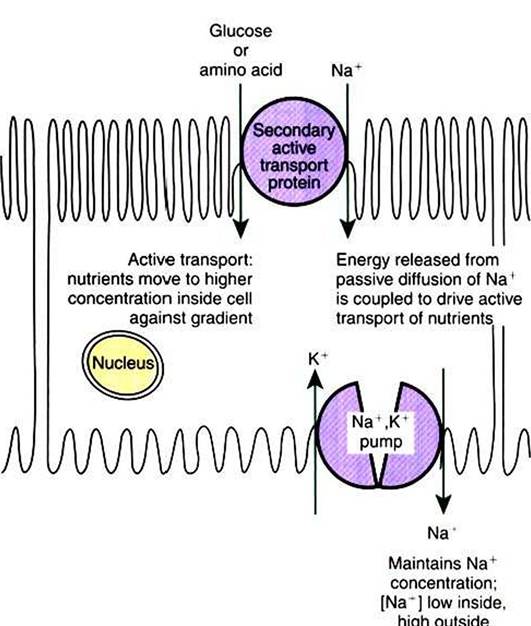
FIGURE 1-9 Secondary active transport as exemplified by uptake of nutrients by gut epithelia. Nutrients such as glucose and amino acids must be actively transported from relatively low concentration in the gut lumen toward higher concentrations within the cells lining the gut.This active transport process uses the concentration gradient of Na* ions set up by Na*,K+-ATPase (see Figure 1-8) as the source of energy for the active transport process.
In other words, the energy released by the passive diffusion of Na* into the cell along its concentration gradient is coupled to the energy-requiring transport of glucose or amino acids against their concentration gradients. Thus the secondary active transport protein both serves a transport function and couples the "downhill" transport of Na* to the "uphill" transport of nutrients.There are many such secondary active transport processes in the body. For example, the same mechanism shown here is used to reabsorb nutrients from blood filtrate in the kidney.That is, the “downhill” flow of Na, from outside the cell to the inside is a spontaneous reaction whose energy can be coupled to some “uphill” reaction (Figure 1 -9). For example, the transport of glucose and many amino acids from the food mass in the small intestine into the cells lining the gut is an active transport process and requires a Na’ concentration gradient. 'Fransport proteins in the plasma membrane of intestinal epithelial cells couple the spontaneous diffusion of Na+ into the cell to the inward, energy-requiring transport of the sugar or amino acids. These nutrients are at higher concentration inside the cell than outside, so they must be actively transported into the cell at the expense of the energy stored in the Na* electrochemical gradient. That is, the energy from the “downhill” diffusion of Na+ into the cell is coupled to the “uphill” transport of the nutrient into the cell. Such active transport coupled to Na’ diffusion across the cell membrane is called secondary active transport because of its dependence on the Na* concentration gradient established by the primary active transport of the Na*,K* pump.
Examples of transport can be identified in a number of ways. Our examples have been instances in which two ions/ molecules must be transported together or not at all, and such transport is called co-transport. Co-transport can involve one process of passive transport (diffusion) with an active transport process, as in the two previous examples; it can involve two active transport processes (e.g., Na*,K1-ATPase) or two diffusion processes. In the first case, the need for co-transport is energetic; the flow of one ion is needed to drive the other. In the two latter cases, the need for co-transport is a restriction based on the binding properties of the transport protein; it cannot bind one without the other. Co-transport proteins that transport both substances in the same direction are called Symports or synιporters. The Na7sugar co-transporter in the gut is a symport. Co-transport proteins that transport the two substances in opposite directions (e.g., Naf, K*-ATPase) are called antiports or antiporters. Parenthetically, proteins that transport only a single ion or molecule are called uniports or uniporters.