Many Membrane Proteins Selectively Facilitate the Transport of lons/Molecules from High to Low Electrochemical Potential
The movement of ions and of medium and large polar molecules requires a protein molecule to serve as a pathway through the obstruction of the phospholipid bilayer. If the movement of the substance is in the natural direction of its electrochemical gradient (movement from high to low), the transport process is called facilitated diffusion.
The membrane proteins mediating this transport process through the phospholipid bilayer are channels or carriers (Figure 1-10). These are distinguished by the extent to which the protein interacts with the transported substance.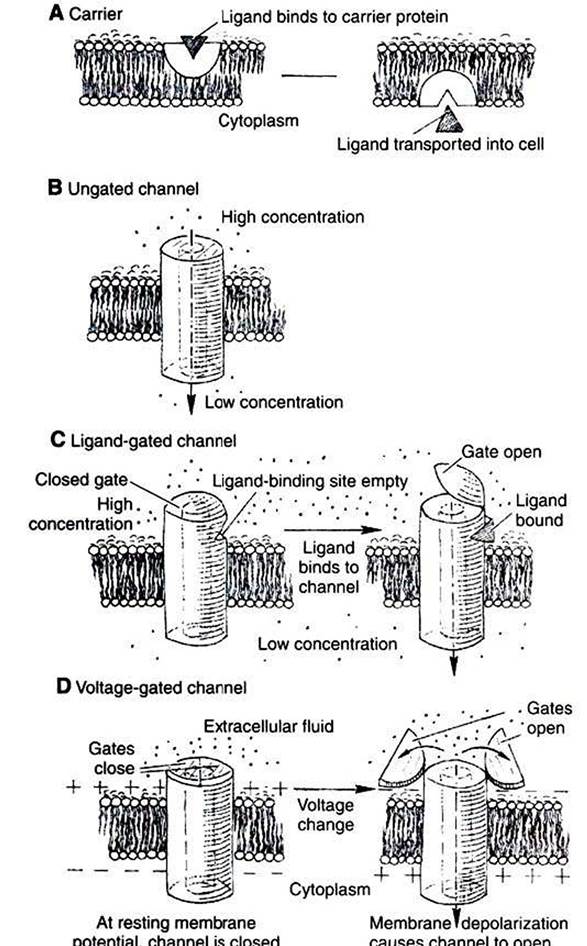
RGURE 1-10 ■ Typesoftransport Proteinsmediating facilitated diffusion. In all cases the ion moves from a region of high potential (shown here as high concentration) to a region of low potential. A, Carriers. In a few cases, material is carried by a transport protein that binds tightly to the material, and the complex moves through the lipid bilayer. B, Leak channels. These channels are thought not to open and close as do gated channels, and thus they support a small but persistent leak of a particular ion through the pore. Although their existence was long postulated, distinct, ungated leak channels have only recently been identified and isolated, as opposed to leaks through normally gated channels. Selectivity of these and other channels is based on the size of the pore and the weak interactions of ions with the atoms lining the pore.
C, Ligand-gated channels. The transport protein again forms a pore through the membrane. In the case of gated channels, access of the ion to the pore is controlled by a gate, a substructure of the transport protein that can open and close the pore. In ligand-gated channels the opening and closing of the gate are controlled by the binding of a ligand to the channel.
D, Voltage-gated channels are similar to ligand-gated channels, except the opening and closing of the gate are controlled by the electrical field around the channel.Carriers bind the transported substance in the Iock- and-key manner, so there is a site-specific binding of the transported substance to the transport protein (Figure 1-10, A). Carrier-mediated transport is typically much slower than channel-mediated diffusion because of the relatively slow binding and unbinding processes. The Na’,K1 pump and the NaVglucose symport are both examples of carriers.
Channels can be thought of as “protein donuts” embedded in the phospholipid bilayer. The hole in the donut is a pore in the membrane through which small ions such as Na+, K*, Ca,, CΓ, and H, are transported. Although most channels transport ions, a class of channels called aqιιaporins comprises channels for water flow. (Although water will flow through a pure lipid bilayer, this transport is too slow for some functions. Kidney cells, for example, are particularly rich in aqua- porins, which are required for the water balance function of the kidney.) For all channels, the pore size and the interaction of the transported material with the amino acid side groups lining the pore allow membrane channels to be selective. Only specific molecules or ions can move through a particular channel. Movement of material through channels is almost as rapid as simple diffusion through a water-filled space of the same area as the channel pore.
The plasma membranes of most cells have passive leaks of ions, particularly K'. These ionic leaks are typically ascribed to leak channels, which are open at all times (Figure 1-10, B). However, most ion channels open or close in response to signals. 'Fhese latter types are called gated channels. The opening and closing of the gates are examples of the allosteric property of proteins. The same signals responsible for allosteric changes in general—ligand binding, phosphorylation, and voltage differences—also control the opening and closing of gated channels, as shown in Figure 1-10.
(Because mechanically gated channels are so poorly understood, these are not discussed here.)Channels that open in response to ligand binding are called ligand-gated channels (Figure 1-10, C). The nicotinic acetylcholine receptor is a ligand-gated channel found in skeletal muscle membrane directly beneath incoming neurons (nerve cells). This channel is found also in the membrane of neurons in autonomic ganglia and in the brain. As the name implies, the nicotinic acetylcholine receptor binds to the drug nicotine and the neurotransmitter acetylcholine. In both cases the channel opens in response to ligand binding.
This nicotinic acetylcholine channel plays a key role in transmitting electrical stimulation from neurons to skeletal muscle cells. Briefly, motor neurons release the neurotransmitter acetylcholine in response to the electrical signal coming down the neuron. This acetylcholine binds to and opens the ligand-gated channel on the skeletal muscle. The influx of Na into the muscle cell initiates an electrical response in the muscle, causing the release of Ca2+ (through gated channels in the endoplasmic reticulum), in turn causing contraction. (This brief account of neuromuscular transmission, presented only to provide orientation to the function of
the acetylcholine channel, is expanded in Chapters 5 and 6.) In the case of the nicotinic acetylcholine receptor/channel, the specific binding and allosteric properties of the protein serve the dual functions of selective transport across the membrane and information reception and transmission to the muscle cell.
Channels that open in response to voltage changes across the membrane are called voltage-gated or voltage-dependent channels (Figure 1-10, D). This type of channel is largely responsible for the neurons’ ability to transmit information along their length and to release neurotransmitter. All voltagegated channels have a range of membrane potentials that cause them to open; this is the channel’s activation range.
'Γhe minimum membrane potential that causes opening is the channel’s threshold. The activation range and threshold vary from channel to channel, depending on the conformation of the protein and the electrical properties of the amino acid side groups that form the gate of the channel. In addition to an open and closed configuration, many voltage-dependent channels have a third conformation, called inactivated. Like the closed configuration, the inactivated conformation prohibits the diffusion of ions through the channel. Unlike the closed configuration, it does not open immediately in response to changes in membrane potential. Inactivation can be regarded as an enforced rest period for the channel. Voltage-dependent channels that do not inactivate have only open and closed conformations, and they take up one or the other conformation, depending on the membrane potential.As previously discussed, any of the functions of proteins can be used to transmit information if a difference in the protein function changed the cell. Gated channels, both ligand and voltage gated, are ideal candidates for information transmission because they change their function: opening and closing, permitting or slopping transport. Indeed, the sole physiological function of the nicotinic acetylcholine receptor/ channel, as described earlier, is the transmission of information: turning the chemical stimulation by the neuron of the muscle into electrical stimulation (see following discussion) of the muscle membrane, leading to muscle contraction.