Most G-Protein-Linked Information Is Sent to the Cytoplasm by "Second Messengers"
As previously noted, the active (heterotrimeric) G protein stimulates an enzyme or ion channel that is associated with the plasma membrane. The ensuing change in ion channel or enzyme function can alter the membrane potential or cause certain molecules/ions to change their concentration in the cytoplasm.
Those ions and molecules that are linked to receptor-ligand binding are called second messengers. A second messenger is an ion or molecule that carries the information within the cytoplasm of a cell in response to a signal on the outside surface of a cell (the first message), such as the binding of a hormone or neurotransmitter, or to an electrical event. Most G-protein-linked information is transduced into the cytoplasm in this manner. One of the major advances in our understanding of the molecular basis of physiological signaling is the realization that there are only a few second-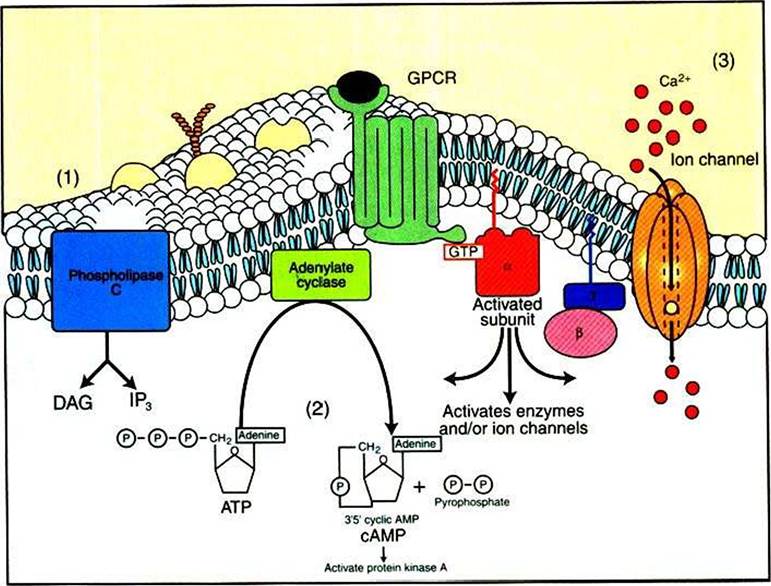
FIGURE 1-16 Activated a subunit of the G protein (GJ can activate enzymes and ion channels, leading to second-messenger signaling within the cytoplasm. Three principal second messengers send the GPCR information to the cytoplasm. These arise from the activation of ion channels and enzymes stimulated by Gu.The second messengers are (1) increases in the concentration of inositol 1,4,5-trisphosphate (IP3) in the cytoplasm and increases in the concentration of diacylglycerol (DAG) in the plasma membrane, both as a result of the breakdown of a rare membrane phospholipid, phosphatidylinositol 4,5-bisphosphate (PIP2) by phospholipase C, another Gu-Stimulated enzyme; (2) changes in the concentration of cyclic AMP (cAMP), a special hydrolytic breakdown product of ATP created by the enzyme adenylyl cyclase, which can be activated or inhibited by α subunits; and (3) changes in Ca2* concentration within the cytoplasm resulting from transport of Ca2* through gated channels stimulated by Gu.
messenger systems within animal cells.
The most important include the following (Figure 1-16):1. Two second messengers, inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG), are produced by G-protein activation of an enzyme phospholipase C (PLC) (see Figure 1-19 and later discussion).
2. Changes in the concentration of cyclic adenosine monophosphate (cAMP).
3. Changes in Ca2+ concentration within the cytoplasm.
Clearly, there are many more CiPCRs than second messengers. This means that several receptor-mediated events are converted into the same intracellular signal. How does the cell sort out this information? Different cells respond differently to the same second-messenger ion/molecule as a result of the specialized function and makeup of that cell (i.e., the differentiated state it achieved during the development of the animal). For example, smooth muscle cells respond differently to activation of muscarinic acetylcholine receptors (see Table 1-2) than do nerve cells because the two cells have different proteins that are responsible for their specialized tasks. However, this is only part of the answer, and the specificity of response to the same second messenger and to activation of similar or identical receptors remains an important open question in physiology.