G-Protein-Coupled Receptors Are the Largest Family (a "Superfamily") of Receptors and Help Regulate Almost All Physiological Processes
It would be difficult to exaggerate the importance and versatility of information processing that begins with a signal molecule binding to G-protein-coupled receptors (GPCRs). There are approximately 900 GPCRs in humans (Table 1-2).
Table 1-2
Partial List of G-Protein-Coupled Receptors (GPCRs)
| ReceptorZreceptor family* | Example of function | Drug ligands |
| α-Adrenergic | Regulates vasculature | Phenylephrine, oxymetazoline |
| P-Adrenergic | Regulates heart and vasculature | Atenolol, propranolol |
| Angiotensin | Principal regulator of blood pressure | Losartan |
| Calcitonin | Regulates bone resorption | t |
| Cannabinoid | Unknown but found widely in brain | Marijuana and derivatives |
| Dopamine | Movement, cognition, and emotions | Chlorpromazine, bromocriptine |
| Frizzled | Regulates proliferation and differentiation, particularly in stem cells | t |
| Gastrin | Regulates acid secretion in stomach | Pentagastrin |
| Glucagon | Regulates “starvation" response | Exendin-4 |
| Histamine | Mediates inflammation and allergy | Diphenhydramine, chlorpheniramine |
| Muscarinic | Secretion of hormones and neurotransmitters | Atropine, carbachol |
| Olfactory | Mediates smell | t |
| Opiod | Mediates analgesia | Morphine, codeine, heroin |
| Opsins | Mediates light transduction in retina | t |
| Prostaglandin | Vasodilation | Sulprostone |
| Serotonint | Regulates gut motility, behavioral arousal, feeding, circadian rhythms | Sumatriptan, ketanserin |
| Vasopressin | Regulates water balance of body | Terlipressin, desmopressin |
*ln most cases, receptor is named for its ligand.
tNone commonly known.tOne member of the serotonin receptor family is not G-protein-coupled.
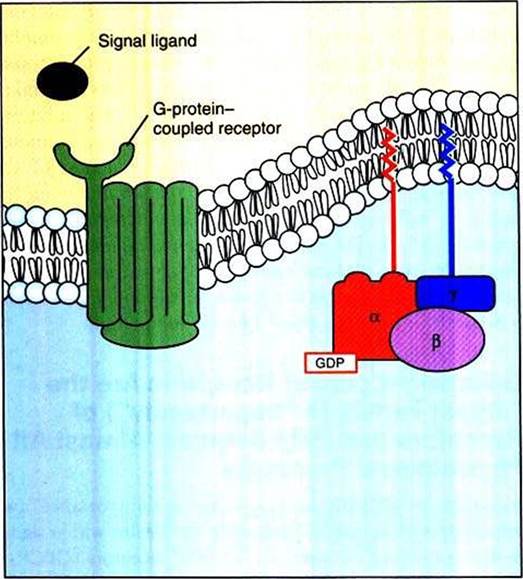
FIGURE 1-14 G-protein-coupled receptor (GPCR) and the heterotrimeric G protein.The hundreds of GPCRs share a similar protein shape, snaking in and out of the membrane seven times.Thus, GPCRs are also called "serpentine receptors" and "heptahelical receptors."These receptors interact with a membrane-associated guanosinetriphosphatase (GTPase) molecule composed of three different polypeptide subunits ("heterotrimeric").The heterotrimeric G protein is not an intrinsic membrane protein, but rather associates with the membrane through lipid tails inserted into the membrane.
There are an even greater number in animals that depend more on olfaction, with about 1300 in rodents, because smell is mediated by different odorants binding to different GPCRs. An estimated 40% to 50% of all commercial drugs act in a GPCR pathway, exemplifying the importance of GPCRs to medicine. All GPCRs share a similar molecular shape; they are an integral membrane protein composed of a single polypeptide chain that passes in and out of the plasma membrane seven times, resembling a snake (Figure I -14). As a result, two other names for GPCRs are seven-transmembrane receptors and serpentine receptors. However, the name GPCRs is more revealing about their mechanism because all also share the same “next step” in their Rube Goldberg signal sequence: they activate a molecular “on-off switch” known as a G protein, so called because it is a guanosinetriphosphatase (GTPase).
GPCRs bind to a particular type of G protein (another of the many “families” of informational proteins), which is a membrane-associated trimeric protein composed of α, β, and γ subunits. Thus, this type of G protein is called the heterotrimeric G protein (“three different subunits”).
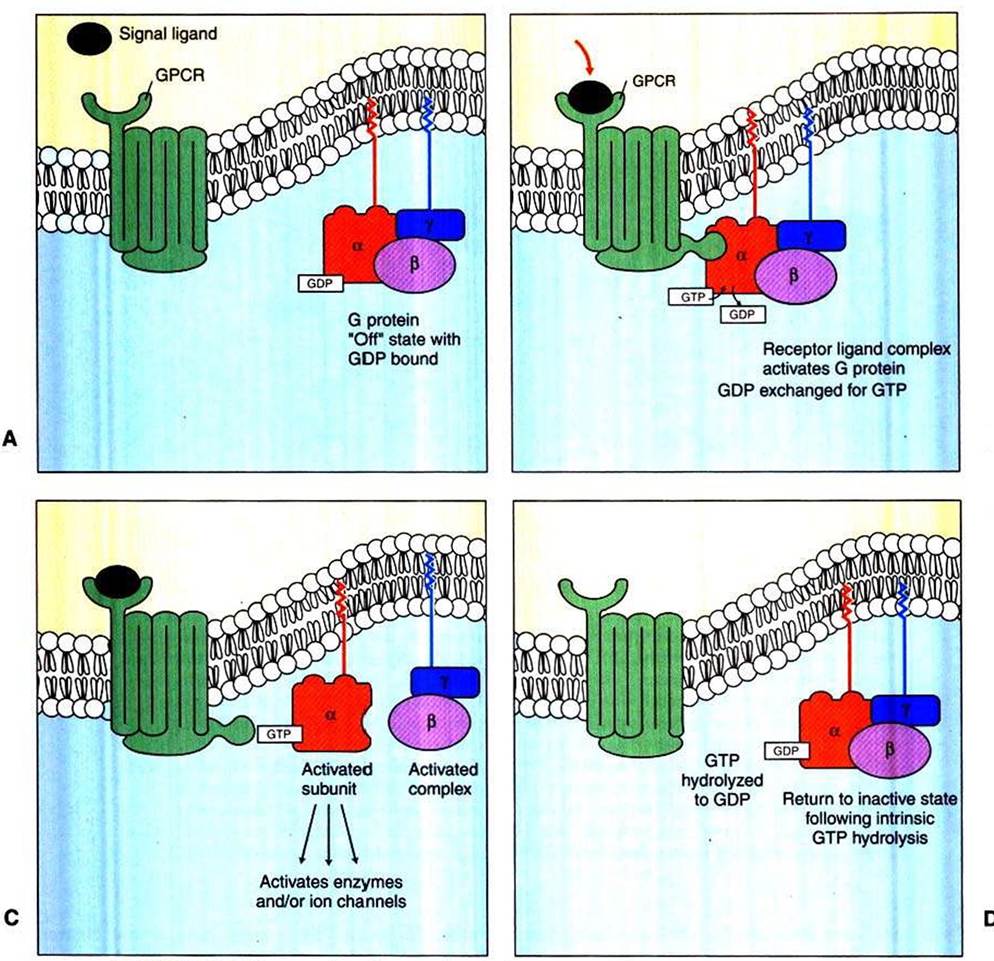
The heterotrimeric G proteins bind directly to the cytoplasmic domain of a GPCR. Although not intrinsic membrane proteins, heterotrimeric G proteins are closely associated with the plasma membrane through lipid molecules that are Posttranslationally added to the subunits and that insert into the lipid bilayer (see Figure l-l4).As noted, G proteins are molecular “on-off switches” that are also GTPases activated by the binding of a signal molecule to its cognate GPCR. That is, in addition to binding to GPCRs, G proteins also bind guanosine triphosphate (GTP) and hydrolyze it to guanosine diphosphate (GDP). The binding and hydrolysis of the GTP to GDP underlie the biochemical mechanism of the on-off switch. In Figure 1-15, A, the unstimulated GPCR does not bind to the heterotrimeric G protein, which is in its “off” state by virtue of the α subunit having GDP and the β and γ subunits bound to it. In Figure 1-15, B, a signal ligand binds to its GPCR, activating the receptor and the G protein. The activation of the G protein takes the form of dissociation of the β∕γ complex from the α subunit, which allows the α subunit to exchange GDP for GTP. The principal “on” activity of the G protein is represented by the Gtt subunit with GTP bound to it. GTP-bound Gu stimulates a variety of enzymes and ion channels that send the signal into the cytoplasm (Figure 1-15, C), as discussed in the next section. However, the G∣iγ complex, once thought to be only an inhibitory Iiictor for the Gu subunit, is now known to activate certain K* channels itself and inhibit certain voltage-dependent Ca2' channels. After stimulating the next element in the signal pathway, the activated GTP-bound Gu subunit returns to an inactive, “off” state as a result of its intrinsic GTPase activity (Figure 1-15, D). That is, the bound GTP is hydrolyzed to GDP, and the Gβγ complex rebinds to the Gu subunit, returning it (and the Gβγ complex) to its inactive state, awaiting the next ligand-receptor-binding event.
As noted earlier, one of the aspects of the Rube Goldberg analogy is that the same conserved types of molecules are often used in many different pathways. Among the many protein “differences that make a difference” to transmit information, one of the most widely used is a GTPase that has on-off states based on whether CiTP or GDP is bound to it. Thus the heterotrimeric G proteins that couple to GPCRs are only one type of GTPase protein acting as an on-off switch in signaling pathways. Most other members of the G-protein (GTPase) superfamily are simpler and resemble the Gu subunit alone. For example, one such class of these “small G proteins,” called Rabs, helps mediate the membrane fusion processes that underlie exocytosis and endocyotosis, discussed earlier. All G proteins share evolutionarily conserved GTP binding and enzymatic hydrolysis sites and a similar on-off mechanism: when GTP is bound, the protein is “on,” and when GDP is bound, the protein is “off.” Chapter 2 discusses a particular small G-protein, Ras, that plays a crucial role in regulating cell division and whose dysfunction plays a major role in cancer. Consequently, G proteins in general are discussed in Chapter 2, and this discussion focuses on signaling mechanisms linked to the heterotrimeric G protein specifically.
B
FIGURE 1-15 Duty cycle of heterotrimeric G-protein, a GTPase that acts as a molecular "on-off switch." See text for further details. A, Unstimulated GPCR is not bound to the heterotrimeric G protein. Bz Signal ligand binds to its GPCRz activating receptor and G protein. Cz GTP-bound Gtf subunit stimulates a variety of enzymes and ion channels that send the signal into the cytoplasm. Dz After stimulating next element in signal pathway, the activated GTP-bound G,f subunit returns to inactive, "off" state because of its intrinsic GTPase activity.