Muscles
Muscular contractions underlie most of the body movements that are grossly visible. Muscle is also used to prevent movement, stabilizing joints to prevent their collapse under a load and maintaining continence of bladder and bowel.
A subsidiary function of the skeletal muscles is to generate heat by shivering, involuntary tremors initiated by exposure to cold.There are three varieties of muscle tissue, but two, the specialized (cardiac) muscle that forms the bulk of the heart and the smooth (visceral) muscle of the blood vessels and viscera (internal organs), are not of present concern. The third variety is generally known as skeletal muscle because the muscles are mostly attached to the bones and induce their movement. Skeletal muscle is also known as striated, somatic, or voluntary muscle, but these terms are less acceptable for one reason or another.
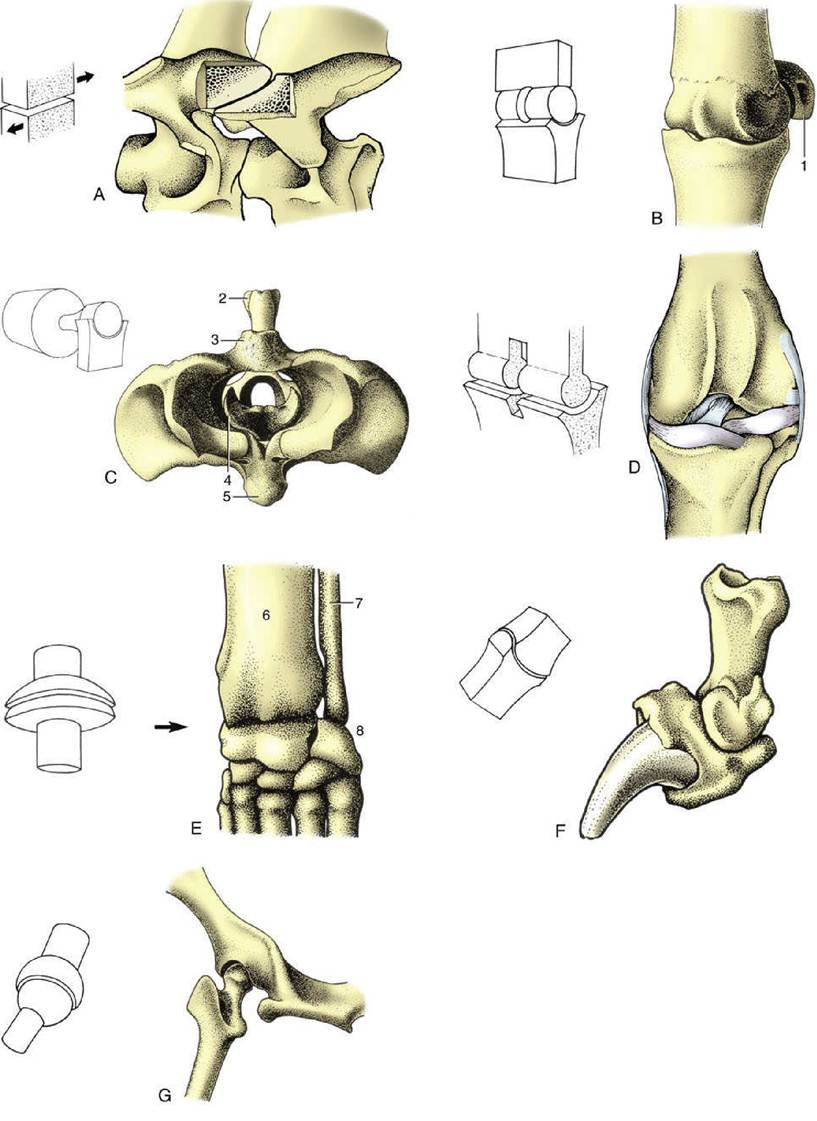
FIG. 1.25 The seven types of synovial joints, with examples. (A) Plane joint: articular processes of equine cervical vertebrae. (B) Hinge joint: equine fetlock (metacarpophalangeal) joint. (C) Pivot joint: bovine atlantoaxial joint (cranial view). (D) Condylar joint: canine femorotibial joint (stifle). (E) Ellipsoidal joint: canine carpus. (F) Saddle joint: canine distal interphalangeal joint. (G) Spheroidal joint: canine hip joint (caudodorsal view). 1, Proximal sesamoid bone; 2, spine of axis; 3, dorsal arch of atlas; 4, dens of axis; 5, ventral arch of atlas; 6, radius; 7, ulna; 8, proximal row of carpal bones.
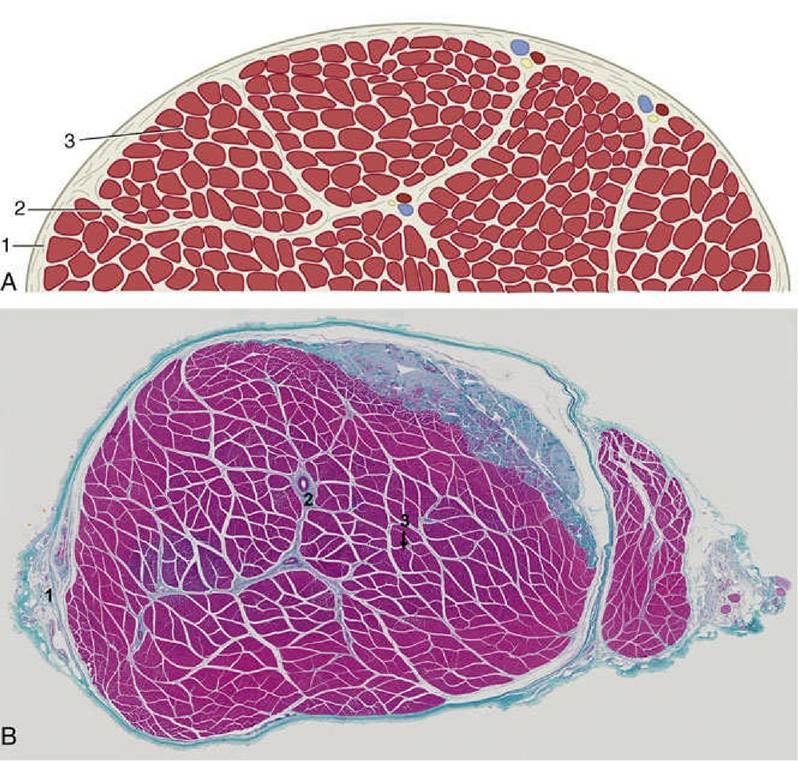
FIG. 1.26 (A) Schematic transection of a skeletal muscle; the fibrous tissue has been emphasized. (B) Cross-section of a skeletal muscle stained with Masson's trichrome stain that shows cellular elements as red and connective tissue as blue.
1, Epimysium; 2, perimysium; 3, endomysium (lightly stained parts in the fascicle).The Organization of Skeletal Muscles
Skeletal muscle is butcher's meat and accounts for about half the weight of an animal carcass (the proportion varies with species, breed, age, sex, and method of husbandry). Each muscle is composed of many cells held together by connective tissue. In comparison with the common run of cell, these muscle cells are giants, varying from about 10 to 100 μm in diameter and being about 5 or 10 cm in length (some are probably much longer). They are visible to the naked eye when teased apart and are also called muscle fibers because of their size and shape. The whole muscle is covered by a dense connective tissue sheet, the epimysium (Fig. 1.26); below this, a looser layer, the perimysium, covers the small bundles (fasciculi) into which the fibers are grouped. Finally, each fiber is provided with its own delicate covering, the endomysium. These connective tissue components merge at each end of the muscle "belly" and continue as the tendons by which the muscle makes its attachment. The amount and quality of the connective tissue partly explain variations in the appearance and in the cooking and table qualities of different cuts of meat (another important factor is the amount of shortening that is allowed by hanging during postmortem rigor).
Variations in Muscle Architecture
There is great variety in the way in which the muscle fibers are arranged within the muscle belly, which can be explained by reference to two principles. The shortening that a muscle may demonstrate on contraction (about 50%) is a function of the length of the component fibers. The power that it may develop is a function of the aggregate of their cross-sectional areas. The greatest displacement, necessarily not the force, is therefore produced by the so-called strap muscle (Fig. 1.27), in which the fibers run parallel to the long axis and throughout the length of the muscle, which is completed by very short tendons of attachment.
Muscles in which the fibers join the tendons at an angle tend to be strong in relation to their bulk because more fibers and a greater total cross section can be accommodated. Although muscles of this sort are powerful, they waste a proportion of their strength and their potential for displacement; only part, corresponding to the cosine of the angle of fiber insertion, is applied along the line of pull. In calculating the power that such muscles develop, one needs to replace the simple "anatomic" cross section with the "physiologic" cross section, which is the complex plane that cuts each component fiber transversely. Muscles with angled fibers can be arranged in several categories of increasing complexity of construction: pennate, bipennate, circumpennate, and multipennate (see Fig. 1.27).
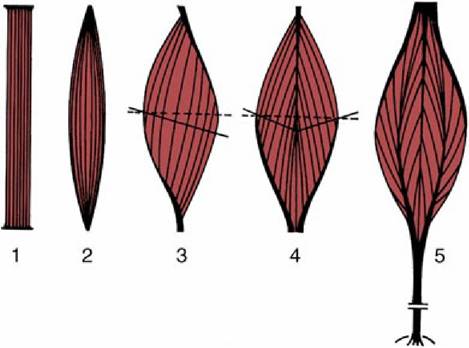
FIG. 1.27 Architecture of skeletal muscles. The broken lines represent the “anatomic,” the solid lines the “physiologic” transverse sections. 1, Strap muscle; 2, spindle-shaped muscle; 3, pennate muscle; 4, bipennate muscle; 5, multipennate muscle.
Many limb muscles have a pennate form and, unlike the strap muscles, have long, cordlike tendons that permit the heavy bellies to be placed close to the trunk; because only the light tendons extend to the digits to operate the joints, less energy is required to swing the limb to and fro. Certain muscles of the body wall form thin flat layers that are continued by broad tendon sheets (distinguished as aponeuroses), an arrangement clearly adapted to supporting the abdominal organs. Other muscles arise by two, three, or four separate heads that join in a common tendon; these arrangements are indicated by inclusion of the descriptive term biceps (two-headed), triceps, or quadriceps, respectively, in the muscles' names.
In another less common variety, two or more fleshy units are separated by intermediate tendonforming digastric (two-bellied) or polygastric units.
Still other muscles are arranged in rings that surround natural orifices, such as the mouth or anus, and act as sphincters to constrict or close the opening. In all these examples the construction of the muscle is clearly adapted to its functions.Paired muscles lying against, or originating from, the midline are separated by a connective tissue strip known as a raphe.
The color of muscles is a reflection of the amount of myoglobin in their fibers. The difference, well exemplified by the pale breast and dark leg muscles of the chicken, is generally regarded as reflecting a pale muscle's adaptation for rapid contraction over a short period and a darker muscle's adaptation for slower but sustained activity. Most muscles are actually composed of two fiber types in varying proportions: fast twitch fibers that rely on glycolytic metabolism predominate in dark (red) muscles, and slow twitch fibers that obtain their energy from aerobic metabolism in pale (white) muscles. There are many other structural and physiologic differences between fibers, and the suggestion that there are only these two, sharply distinguished varieties, although convenient, is a misleading simplification.
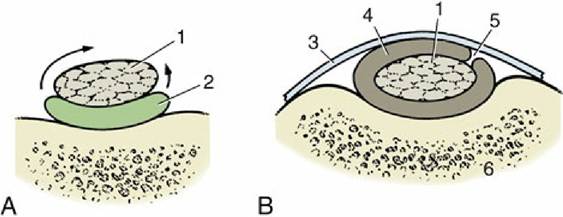
FIG. 1.28 Sections of a synovial bursa (A) and a tendon sheath (B). The bursa permits frictionless movement of a tendon (1) over bone, and the sheath permits movement of a tendon over bone and under a retinaculum. The arrows show that a tendon sheath may be regarded as a large bursa that has wrapped around a tendon. 2, Bursa; 3, retinaculum; 4, tendon sheath; 5, mesotendon, through which blood vessels reach the tendon; 6, bone.
Tendons
Muscles always attach by means of connective tissue tendons. Sometimes the tendons may be so short as to create the illusion that a muscle directly attaches to the bone. Tendons consist almost entirely of collagen bundles in regular arrangement, and they possess great tensile strength.
Indeed, excessive tension is more likely to rupture the muscle belly or to detach a fragment of bone at the insertion than to disrupt the tendon itself. The elastic nature of tendons allows them to absorb and store energy to generate recoil that aids in locomotion. Also, a good fraction of the metabolic work performed by many muscles is devoted to stretching tendons so that the stored energy can later be released.Although they are tough, tendons may be damaged by excessive pressure or friction, particularly when they change direction over bony prominences or are shifted over hard tissues. One form of the protection that they develop in such places, local chondrification or ossification (sesamoid bones), has been mentioned. An alternative is provided by the development of fluid-filled cushions at the danger sites. If only one aspect of the tendon is at risk, a bag (bursa synovialis) may be interposed on that side (Fig. 1.28A); if a greater part of the circumference is vulnerable, the cushion wraps around the tendon, enclosing it within a tendon sheath (vagina synovialis; Fig. 1.28B). The walls of these bursae and sheaths and the fluid they contain resemble the similar components of synovial joints. When the tendon moves, it is the lubricated synovial layers that rub together.
Inflammation of synovial bursae and sheaths is common, and it is necessary to know their positions and extents; however, this is not difficult because they occur precisely where they can be seen to be required.
Blood and Nerve Supplies of Muscles
Muscles receive a relatively generous blood supply from neighboring arteries. Sometimes, a single artery enters the muscle belly, and then the well-being of the muscle clearly depends on the integrity of that artery. Often, two or more arteries enter separately and connect with each other within the muscle. Unfortunately, these connections (anastomoses) are not always sufficient to allow the muscle to survive unscathed an interruption to one of its sources of supply.
The intramuscular arteries ramify within the perimysium to open into capillaries that follow the endomysial sheaths of individual fibers.The veins are satellite to the arteries. Normal activity, during which only a fraction of the muscle fibers contract, probably promotes the circulation within the muscle by massaging the capillaries and smaller veins. Mass contractions squeeze these vessels from all sides, stopping the circulation, and are likely to be harmful if sustained.
Tendons have low metabolic needs, are poorly vascularized, and do not hemorrhage when cut. These features, initially apparent advantages, have their adverse side: damaged tendons are inevitably slow to heal. Lymphatic vessels are found within the larger connective tissue tracts of the muscle belly.
Most muscles are supplied by a single nerve, but those of the trunk that are formed from several somites (p. 28) retain multiple innervation. The nerve that enters a muscle, generally in company with the principal vessels, ramifies within the connective tissue septa. It consists of fibers of several types: large alpha motor fibers supply the muscle fibers of the main mass; smaller gamma motor fibers supply modified muscle cells within the muscle spindles buried in the muscle; nonmyelinated vasomotor fibers supply blood vessels; and sensory fibers supply the spindles, tendon organs, and other receptors. The ratio of motor to sensory fibers varies considerably and is one among many complications in the determination of motor unit size.
The motor neurons that supply a particular muscle are roughly grouped within the ventral horns of gray matter in the spinal cord (or within motor nuclei of the brainstem). The axon from each neuron branches repeatedly in its passage, both within the nerve trunk and within the intermuscular septa, and ultimately ends in the motor end plates of several or many muscle fibers. The single neuron, as well as the (alpha) fibers it supplies, is known as the motor unit, an important concept because it is the physiologic unit of muscular contraction. It is these groups and not individual fibers that are called up for or discharged from service when a muscle varies the force of its contraction. The muscle fibers belonging to a unit are intermingled with those of other units and do not correspond with any readily identifiable portion of the muscle—they do not correspond with the fasciculi, as one might suppose. The fibers constituting a motor unit are invariably of a uniform type.
In the human species, the numbers of fibers within a unit varies, numbering about 5 to 10 in the muscles that move the eyeball, around 200 in the muscles of the fingers, and approximately 2000 in the muscles of the limbs. The exact figures are not important, but the trend is: the muscles with the smallest units are those capable of the most delicate adjustment. Motor unit size is determined from the innervation ratio, the ratio between the numbers of fibers within a muscle and the numbers of motor neurons that supply it.
Muscle Actions
When a muscle is activated, its fibers attempt to shorten. When shortening occurs, the tension in the muscle may increase, stay the same, or decrease, according to circumstances. When external forces prevent the muscle from shortening, the tension within it increases; such activity is said to be isometric.
The usual activity of most muscles involves changes in the angle of the joint(s) bridged by that muscle. The musculoskeletal system thus operates as a system of levers in which the joints act as fulcrums. The mechanical advantages of the arrangement depend on the positions (relative to the fulcrum) of the muscle attachment and the application of the load (Fig. 1.29). Although a muscle attaching close to a fulcrum is less powerful than a comparable muscle attaching at a greater distance, it produces its effect more rapidly; the requirements of speed and power thus conflict. When several muscles are available to move a joint in a particular way, the attachments of some make them more suited to getting the movement started, whereas the attachments of others make them more suited to carrying the movement through to completion.
Biarticular or polyarticular muscles (those that cross two or several joints, respectively) may be incapable of shortening sufficiently to produce the full range of movement at both or all of the relevant joints at the same time. Such muscles are said to be actively insufficient.
Any muscle that produces a certain effect may be termed an agonist or prime mover; a muscle capable of actively opposing that movement is termed an antagonist. Clearly, these terms have force only in relation to a specified movement. Thus, in flexion of the elbow, the brachialis muscle that produces the movement is the agonist, and the triceps brachii that opposes the movement is the antagonist; in extension of the same joint, however, the triceps muscle is the agonist, and the brachialis muscle the antagonist. Other muscles may neither facilitate nor directly oppose a movement but may modify the action of the agonist, perhaps by eliminating an unwanted side effect. Such muscles are known as synergists. When muscles are employed to stabilize joints rather than to promote their movement, they are known as fixators. Fixation or stabilization of a joint often involves the co-contraction of muscles that oppose each other when the joint is moved.
The terms origin and insertion have been left undefined until now. Conventionally, origin denotes the more proximal or central attachment and insertion the more distal or peripheral attachment. Although it is true that in their common employment most muscles draw the insertion toward the origin, the vast majority are able to shorten toward either end. Which attachment will maintain its position and which will be drawn toward the other depend on external circumstances. These circumstances must always be taken into account when one is considering the possible actions of a muscle.
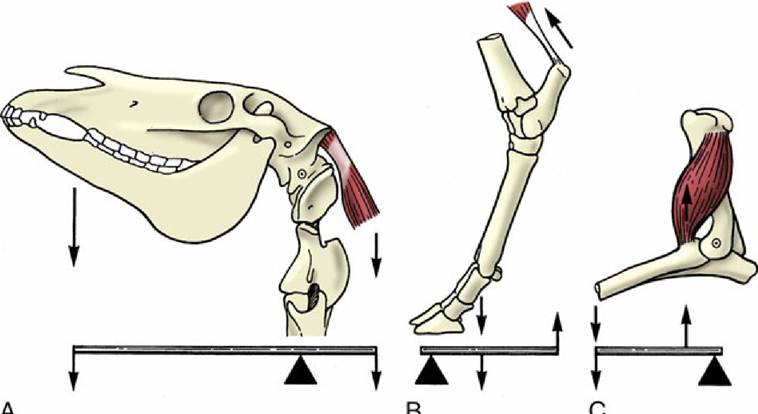
FIG. 1.29 The action of muscles on the skeleton can be compared to different lever systems. (A) Support of the head by dorsal neck muscles. (B) Extension of the hock joint. (C) Flexion of the elbow joint.

FIG. 1.30 Record (electromyogram) of electrical activity during muscle contraction.
The attachments of muscles in relation to the axis (axes) of the joint(s) permit deductions about their functions. If these deductions are sound, they indicate what a muscle can do but not how it is habitually used in life. Direct stimulation of a muscle or of its nerve shows what that muscle can do when it acts alone. It does not, however, show how the muscle is used naturally, because often, several alternative muscles are available to perform a given movement but not all are normally used.
The most elegant technique for studying muscle actions is electromyography, the registration of the electrical activity that accompanies muscular contraction. In this method, electrodes are placed over or inserted into the muscles to time the activity and crudely quantify its intensity (Fig. 1.30). The use of this technique has upset many long-held beliefs concerning the actions and use of the muscles of humans; much remains to be investigated where domestic animals are concerned. Even this method demands that caution be used in interpretation of findings. It shows when a muscle is active but leaves the experimenter to interpret the activity as agonistic, antagonistic, or a mere adjustment to the alteration in joint angle brought about by other forces.