NERVOUS SYSTEM
The ferret brain is about 36 mm long by 24 mm wide and has a normal mammalian structure (Lawes & Andrews 1998). The special senses: hearing, sight, and smell are of interest in relation to conditions and diseases.
The visual and auditory systems mature after birth so that kittens open their eyes at 4-5 weeks and can hear at 32 days.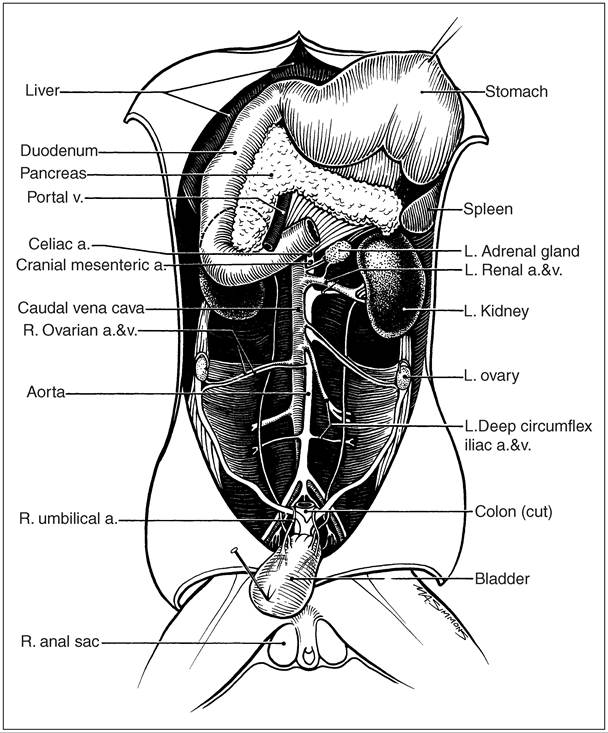
Figure 12.25 • The ferret pancreas, spleen, and ovaries. (Courtesy of Howard Evans.)
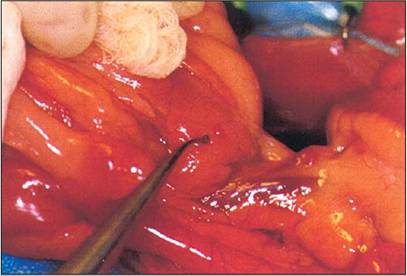
Figure 12.26 • Insulinoma in a ferret. The ring tip microforceps are pointing at a 2 mm nodule typical of insulinoma. Slide courtesy of Prof. Avery Bennett.
in the 4-15 kHz range, while 32-day-old kittens respond to 1-6 kHz but at a higher threshold (Whary & Andrews 1998). Adult frequency response patterns have been recorded at 39 and 42 days. Interestingly, neonates put out distress cries of 100 kHz. The jills with litters respond to all calls above 16 kHz, but this phenomenon is not seen with non-lactating jills presented with neonates.
The question of deafness in ferrets can be a worry but usually, being hunters, ferrets have acute hearing and a keen sense of smell while their sight is more adapted to their nocturnal habit. A Western Australian black-eyed white ferret jill which repeatedly ignored a crying kitten has been recorded as deaf by Amy Flemming (personal communication 1999). It had the Waardenburg syndrome, a dominant genetic fault. Hearing can be checked in ferret kittens at 32 days by their response to a loud clap.
Senses
Hearing
The ferret auditory system is physiologically similar to that of the cat, but it is thought auditory response in the ferret is more primitive. Adult ferrets usually respond to sounds
Anatomy of the ear
The external ear consists of a pinna and an external ear canal, which differs markedly in shape from that in the dog and cat (Lewington 1990) (Fig.
12.27). The ferret pinna is set close to the head and half-moon shaped, some 2 cm wide in adult hobs and pointing forward, as is compatible with a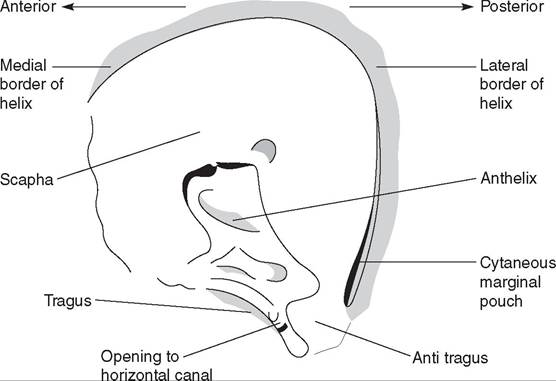
Figure 12.27 • Ferret external ear. (By Debbie Squance.)
hunting animal. Some ferrets have a flatter, wider pinna, large compared to the head size, which is possibly a genetic fault.
There is no distinct tubular ear canal as in the dog and cat. The whole ear canal is protected by a screen of fine hairs around the anterior margin. Interestingly, the lateral margin of the ear canal has a more pronounced recession, intertragic incisure-like, which is more tube-shaped than seen in the cat and some 5 mm long. It gives the impression of a miniature aural resection. Almost opposite, medial to the base of the intertragic incisure, is the opening of the horizontal canal, which passes anteriomedially to the tympanic membrane. There is hardly any depth to it. Whether the shape of the ferret external ear makes it prone to otitis has not been shown.
The middle and inner ear is comparable to that of the dog in general structure (Figs 12.28 and 12.29).
The ear pinna can be the site of fight wounds and advanced cases of Sarcoptes scabies var. canis, which spread from the feet in ferrets. The external ear can be infected with Otodectes cynotis, as in dogs and cats. The inner ear may show idiopathic otitis internal (IOI) with head tilting, and loss of balance due to extensive middle ear infections or blood-borne pathogens (Lewington 2003d).
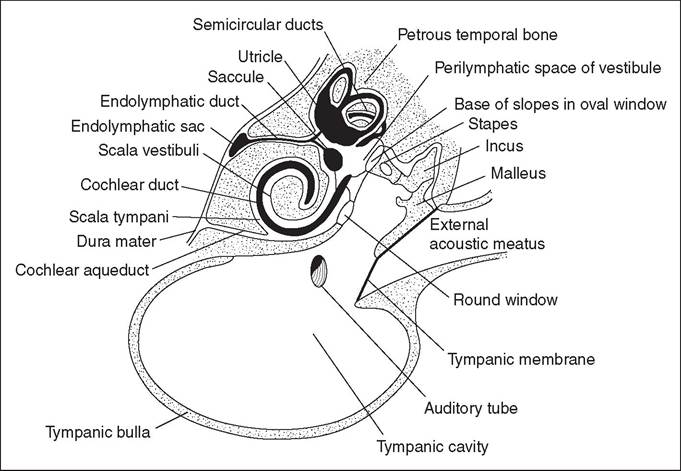
Figure 12.28 • Diagram of the structure of the middle and inner ear of the dog. From Getty, R., Foust, H. L., & Presley et al. (1956) Macroscopic anatomy of the ear of the dog. American Journal of Veterinary Research 17, 364-375, with permission.
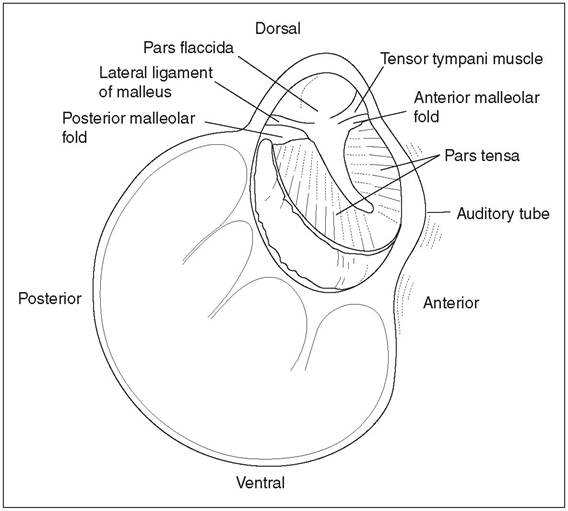
Figure 12.29 • Diagram of the medial face of the lateral wall of the left tympanic cavity of the dog.
From Spreull, J. S. A. (1976) Otitis media in the dog. In R. W. Kirk (ed.), Current Veterinary Therapy V. Philadelphia: WB Saunders, with permission.Vision
The ferret's eyesight is poor in comparison to its senses of smell and hearing; it is adapted to nocturnal life, like its polecat ancestor (Lavers & Clapperton 1990). Studies of the ferret visual system have been orientated to such problems as the influence of photoperiods on reproduction, albinism, and its effects on ocular development, and various studies on the ferret neuro-opthalmic development and basic investigation relevant to human eye problem research (Miller 1997). The visual systems are studied utilizing the very immature state of neonate ferret visual systems (Jackson & Hickley 1995).
Clinical examination of the ferret eye involves normal cat and dog tests except for the Schirmer tear test. The eye can be examined using a magnifying loupe and, for internal eye examination, an opthalmoscope in a dark room. Indirect ophthalmology plus a 30 or 40 diopter (D) condensing lens can be used. Eye specialists use a slit lamp biomicroscope.
Comparative studies of sight in mustelids suggest that the short pointed face of a weasel allows both binocular vision forward and a wide arc of monocular vision on each side (King 1989). One author suggests ferrets have mostly monocular but very little binocular vision (Kircher 1997). Another is of the opinion that from their facial conformation there is no difference in range of vision between weasels, stoats, and ferrets (King 1989).
The eye that sees well in dim light achieves this ability at the expense of acuity in bright light (King 1989). Thus, most carnivores see movements rather than pictures but they can be sharp-eyed on focusing in on prey. So, waving a finger in front of a ferret in daytime can be dangerous: the ferret sees and responds to moving objects at speeds of 25-45 cm/s; for example, a mouse in a hurry! (Miller 1997).
The ferret's ability to see objects clearly depends on the optical properties of the eye, which involves retinal sensitivity and optical processing of visual images in the central nervous system. The retinal activity is usually the controlling factor.
Optical abnormalities can occur in the ferret eye and the optical quality of this special sense organ is considered moderate. In detail the ferret appears to be 6.8 D (range 5-8 D) hyperopic or farsighted. Of 12 ferrets in one study 4 revealed astigmatism of 0.5 D or greater. Additional faults can be found in the refraction between the two eyes because of the large spherical lenses. This may show up in dim light when the pupil is dilated (Miller 1997). On the question of perception of form, work has been done on the weasel, which can appreciate different letters in form discrimination tests (Herter 1939) to get a food reward, or not, from differently labelled boxes (King 1989). In the ferret image brightness might be a factor as objects brighter than the background stimulate a more effective pre-catching behavior than darker images (Miller 1997).Color vision in ferrets is possible. Experiments with weasels and stoats (Gewalt 1959; Herter 1939), which have rods and cones in ratios typical of diurnal animals, have shown that the stoat can see at least red and possibly also yellow, green, and blue. The ferret has not been studied but its retina is similar to the cat's in its density of rods and cones, so there is a suggestion that ferrets can detect color (Miller 1997).
258
In any case the detection of color is of no real advantage in nocturnal animals.
Anatomy of the eye
The ferret has a well-developed nictitating membrane which can be used for ulcer treatment as a third eyelid flap (Lewington 1994). The cornea is relatively large in relation to the eye structure and the lens is nearly spherical and placed back in the posterior eye chamber (Kern 1989). The pupil is a horizontal elliptical slit, not vertical like the cat, and more efficiently protects the eye from strong sunlight.
Retina
The retina is similar in form and vascular pattern to that found in the dog (Miller 1997). The tapetum is well defined with the 7-10 cell layer having a high zinc and cysteine content; the myelinated optic disk is relatively small (Wen et al.
1985). A study of the tapetum lucidum established that the structure is the same in both pigmented and albino ferrets (Tjalve & Frank 1994). The receptor cells in the retina, cones, and rods, differ in function; the cones perceive bright light and colors and are found in large numbers in diurnal species whereas rods are particularly sensitive to low-intensity light and are numerous in the eyes of nocturnal animals (King 1989). Thus the ferret retina has a high proportion of rods in the photoreceptor layer of the ferret eye. The rods predominate to the cones in the ratio 50-60:1 (Jackson & Hickley 1995).CLINICAL NOTE
The ferret, like other mustelids, having the combination of a large cornea, spherical lens, and a tapetum lucidum highly receptive to dim light, is well-adapted to nocturnal living.
The tapetum gives the “eye-shine” glow to nocturnal animals caught in a torch beam.
Photoperiod
The ferret eye, in association with the visual nervous system linked to the pineal gland, is involved with reproduction and also with endocrine problems, for example, the jill ferret estrous cycle is related to photoperiod length. The cycle will begin with extended photoperiods in spring and can be adapted to increasing or decreasing the number of litters per year. A blind ferret will not start estrus. Increasing photoperiods stimulate the hob to become sexually active. It is theorized that an artificial regimen of extended photoperiods may cause adrenal gland neoplasia in the sterilized ferret due to the failure of melatonin production in the continuously light-stimulated pineal body.
Olfaction
The ferret's sense of smell ranks in importance with its sense of hearing (Lavers & Clapperton 1990). Marking territory and scenting another animal's presence and path is common to many mammals. Mustelids are persistent in following the scent of prey.
Nasal anatomy
The pointed face of the ferret has a tapering nasal organ (see Fig. 12.9). The nasal cavity is formed by the maxilla and nasal bones dorsally and laterally, with the maxillary and palatine bones supplying the cavity floor, similar in shape to the long-nosed dog.
The bony nasal aperture is composed of two symmetrical halves (Evans & de Lahunta 1971).The ethmoid bone complex is located between the brain case and the facial part of the skull. It consists of the ethmoid labyrinth, the cribriform plate, and the median bony plate of the nasal septum. The ethmoid labyrinth is composed of many delicate scrolls, as in the dog, that attach to the cribriform plate and occupy the fundus of the nasal cavity. Bony scrolls, the conchae, project into each nasal fossa and with their mucosa act as baffles to warm and clean inspired air. The nasal cavity is therefore one of the first lines of defense against pathogenic invasions. On the other hand it can supply a vascular environment for pathogens to grow and spread if they overcome the body's defenses. The conchae are divided into a small dorsal and large ventral portion, as in the dog. The olfactory mucosa lining the conchae that house the olfactory nerve cells allow infection to invade the brain via the cribriform plate of the ethmoid complex. The ethmoid plate in the ferret does not completely pass ventrally to the cartilage of the nasal septum to divide the nasal fossa as it does in the dog. Thus it is more difficult to pass a nasal tube in the ferret, but it is not impossible (Lucas 2000) (Fig. 12.30).
Clinically, the nasal cavity is an entry point for viral infections, such as canine distemper and human influenza, and a gateway for infection of the lungs by a variety of organisms leading to pneumonia. The closeness of the cribriform plate to the brain meninges can be an entry to such pathogenic fungi as Cryptococcus neoformans, leading to acute meningitis in the ferret (Lewington 1997).