Overview of motor and sensory systems
Key points
■ The motor system maintains static and dynamic posture, controls voluntary movement, gait and visceral motor function. It can be divided into two components, upper motor neurons (UMN) and lower motor neurons (LMN).
■ The neuromuscular junction connects the electrical activity of the LMN to the depolarisation of the muscle via chemical messengers.
■ Planning and direction of voluntary movement is centred in the forebrain. The forebrain and the cerebellum communicate with UMNs in the motor cortex and brainstem. The UMNs interact with LMNs in the brainstem and spinal cord.
■ Sensory receptors can be classified based on morphological criteria, the type of sensory modality to which they are receptive and by the type of the animal’s environment they are sensing.
■ Sensory receptors are distributed throughout the body and head. Their axons project into the CNS, making local connections with LMNs for reflex function. They also project to centres in the brainstem and forebrain for integration.
Overview of motor (output) systems
The functions of the motor system are (a) to maintain posture and provide a stable platform for movement, (b) voluntary movement and locomotion, and (c) visceral motor function (see Chapter 5).
Upper motor neurons, lower motor neurons and the neuromuscular junction
The motor system can be divided into two main components - upper motor neurons (UMNs) and lower motor neurons (LMNs). The UMNs are completely contained within the CNS, thus an alternative name for them could be ‘central motor neurons’. The UMN cell body is located in a motor nucleus of the brainstem or the motor cortex of the forebrain. Their axons connect to LMNs either by synapsing on them directly or indirectly, via interneurons. The LMNs have their cell body in the CNS (brainstem or spinal cord).
Their axons project into the PNS, via cranial or spinal nerves, to connect with striated or smooth muscle at the neuromuscular junction; thus an alternative name for them could be ‘peripheral motor neurons’. Note that while the soma of the LMN connecting to striated muscle lies in the CNS, its long axon is located in the PNS. The ANS has two LMNs in series, with the cell body of the second LMN located in a peripheral ganglion (see Chapter 12). The UMNs, along with other management components of the motor system, organise and direct the activity of the LMN. When stimulated LMNs induce muscle contraction. Thus the UMNs are part of a motor management system, while the LMNs with their associated muscles are analogous to the workers.The neuromuscular junction (NMJ) (see Fig. 1.4) connects the electrical activity of the LMN to the depolarisation of the muscle, using chemical messengers, such as acetylcholine (ACh). Even in the absence of action potentials, acetylcholine is continually being released at the NMJ causing miniature depolarisations of the muscle end plate. These miniature end plate potentials result in end plate noise that can be detected electromyographically using needle electrodes. Release of ACh also contributes trophic support to the muscle fibres, which when lost, as occurs when the LMN is destroyed, is associated with rapid, severe, neurogenic atrophy of the muscle fibres (see Chapter 5). When a volley of action potentials arrives at the motor end plate, large amounts of ACh are released and the muscle is stimulated to contract proportionally.
There are two basic activities of striated muscles in the body and limbs, extension and flexion of joints. Extensor muscles are antigravity muscles as they permit the animal to resist the effects of gravity. Extensor muscles increase the joint angle. For the thoracolumbar vertebral column, flexor (hypaxial) muscles work in conjunction with extensor (epaxial) muscles to resist the effects of gravity on the trunk.
Antigravity muscle activity has to dominate flexor muscle activity, otherwise the animal would be recumbent. However, in the normal animal, extensor dominance is modulated by the action of inhibitory UMNs synapsing on the LMN supplying extensor muscles. Thus the overall descending UMN activity in the normal animal is inhibitory to antigravity extensor muscles. Damage to these inhibitory UMN pathways results in extensor dominance and spasticity (Fig. 4.2).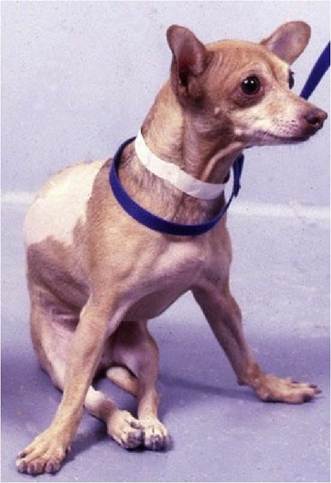
Fig. 4.2 Extensor dominance in the pelvic limbs of a dog due to a lesion disrupting UMN tracts in the caudal thoracic spinal cord. The thoracic limbs are also in increased extension (braced) to carry the weight that is no longer being borne by the pelvic limbs and extensor muscles of the caudal half of the trunk.
Overview of sensory (input) systems
Sensory receptors can be classified in several ways, based on morphological criteria (e.g. encapsulated or nonencapsulated) or the type of sensory modality type to which they are receptive. They can also be classified based on what part of the animal’s environment they are sensing. This could be the external environment (exteroceptors), or the internal environment (interoceptors) or receptors that specialise in stimuli used in spatial orientation of the animal (proprioceptors). Examples of exteroception include vision, audition, olfaction, gustation and touch. Examples of interoceptors include receptors in the viscera sensitive to stretch, pH, and chemical changes such as CO2. Proprioceptors are located throughout the body and head, and in the vestibular apparatus of inner ears. They include muscle spindles (stretch receptors), Golgi tendon organs, joint receptors, tactile/pressure receptors, especially of the feet, and hair cells of the inner ear.
Worker and management systems
Motor systems
The workers comprise the LMNs in which the motor neuron connects directly with the muscle.
Superimposed on the workers are the various levels of management. Sitting in a role analogous to junior management is interneuronal circuitry that connects sensory and motor nerves and different groups of LMNs so they can function as integrated, coordinated worker systems. Junior management is located near the LMN cell bodies in the spinal cord or brainstem and connects to the LMNs of the spinal or cranial nerves, respectively. It receives direction from senior management.Senior management comprises the traditional UMN tracts that, in domestic mammals, originate primarily in the brainstem and in the motor cortex of the cerebral hemispheres.
Executive management has its head office mainly in the cerebral cortex, but it also has diverse connections and feedback loops throughout the brain including with the cerebellum and basal nuclei.
Functionally, executive management holds primary responsibility for planning and directing voluntary and complex movement. It talks to senior management. Senior management receives instructions from the executive, has feedback loops with some areas of the brain, especially the cerebellum, and directs junior management (interneurons) in the brainstem and spinal cord. Senior management can also interact with workers (LMN) directly. When the UMNs fire, they can stimulate, or inhibit, the activity of the LMN. Junior management is like the foreman on the workshop floor, listening to input from both the workers and the managers, and coordinating the outputs of worker groups to produce the specific patterns of movements that are required for actions like locomotion.
Sensory systems
For the sensory systems, the workers are the receptors distributed throughout the body (extero-, intero- and proprioceptors) with their axons projecting via cranial and spinal nerves into the CNS.
Once inside the CNS, the fibres make connections with local circuitry (junior management) for reflex function. For spinal nerves, these connections are made primarily with interneurons in the dorsal horn, which then ultimately connect with LMNs in the ventral horn.
For example, a noxious stimulus applied to the foot travels via the spinal nerves to the dorsal horn. From there it connects via interneurons to LMNs in the ventral horn of several spinal cord segments stimulating LMNs supplying limb flexor muscles and the foot is withdrawn from the stimulus. Cranial nerves convey sensory input to brainstem cranial nerve nuclei. This input connects via interneurons to LMN in other brainstem nuclei. For example, in the palpebral reflex, input to the trigeminal nucleus connects via interneurons to the facial nucleus so that touch around the palpebrum triggers reflex blinking.Connections from sensory spinal nerves can also travel cranially in the cord to synapse in brainstem nuclei and make connections with UMNs (senior management of the motor system). Sensory information is also projected directly to primary sensory receiving areas in the cerebellum (for subconscious proprioception) and the somatosensory cortex of the cerebrum. Input from cranial nerves is projected to the forebrain where it is received in auditory, somatosensory, olfactory and visual cortices of the cerebrum. Input from the vestibular system (head proprioception) is also projected to the cerebellum via CN VIII. These primary receiving areas are like the senior management for the sensory system.
Sensory input that is received in the primary sensory cortices is projected to nearby association areas where it is processed, ranked in terms of importance and, from there, integrated with other information. The association and integration areas are the equivalent of the executive management for the sensory systems. For example, visual input via cranial nerve II is received in the visual cortex in the occipital lobes. The executive system for visual input will process that information in the nearby visual association area and link it with other processed sensory inputs (e.g. touch, audition), stored information (memory), emotion-generating areas of the brain (limbic system) and information from other integrating and coordinating centres such as the cerebellum and basal nuclei. The locations of the main functional regions of the cortex are comparable in all mammals, but their extent varies: for example in the carnivore the primary sensory and motor areas and olfactory cortex comprise 80% of the cortical surface whereas in primates the same zones make up less than 20%. The relationship is reversed for the associative cortex in these two groups of animals, while rodents and lagomorphs have almost no associative functions at all (see Fig. 4.16).
The sensory executive areas will also link with the executive motor areas for planning of voluntary movement. Thus sensory systems can modify motor output directly at the local level via reflexes utilising junior management, or indirectly via connections to the motor system hierarchy at senior and executive management levels.