THEORIES OF AGING
Aging is a multi-factorial process. Most hypotheses on the underlying mechanisms of the aging process involve the deterioration of the maintenance of homeostatic metabolic, inflammatory, and/or redox processes in cells and tissues.
25.5.1 General Wear-and-Tear Theory
Dr. August Weismann, a German biologist, first introduced this theory in 1882. He believed that the body and its cells were damaged by overuse and abuse. The organs and skin are worn down by toxins in the diet and in the environment. Wear and tear are not confined to the organs; it also takes place at the cellular level. As one gets older, small traumas to the body build up. Point mutations increase in number, and the efficiencies of the enzymes encoded by our genes decrease. Moreover, if a mutation occurred in a part of the protein synthetic apparatus, the cell would make a large percentage of faulty proteins. If mutations arose in the DNA-synthesizing enzymes, the number of mutations would be expected to increase markedly. Faulty DNA polymerases in senescent cells have been evident. Likewise, DNA repair may be important in preventing senescence, and species whose members' cells have more efficient DNA repair enzymes live longer. Moreover, genetic defects in DNA repair enzymes can produce premature aging syndromes inhumans.
25.5.2 Neuroendocrine Theory
This theory was described in the year 1954 by Dr. Vladimir Dilman, a Russian scientist. The neuroendocrine theory of aging states that ”the effectiveness of the body’s homeostatic adjustments declines with aging - leading to the failure of adaptive mechanisms, aging and death.” This theory has also been referred to as the aging clock theory. Consistent with this theory, the hypothalamic-pituitary-adrenal (HPA) axis, the main regulatory system controlling homeostasis in humans, loses efficiency with aging. The HPA system works by the interplay of various hormonal signals that initiate reactions in target tissues coupled with a negative feedback mechanism to allow for fine control of body functions, such as blood pressure, fluid and electrolyte levels, and body temperature.
This theory proposes that aging is due to changes in neural and endocrine functions that are crucial for 1. Coordinating communication and responsiveness of all body systems with the external environment, 2. Programming physiological responses to environmental stimuli, and 3. Maintaining an optimal functional state for reproduction and survival while responding to environmental demands. Hormones are vital for repairing and regulating bodily functions, and when aging causes a drop in hormone production, it causes a decline in the body’s ability to repair and regulate itself as well. Thus, hormone replacement therapy, a frequent component of any anti-aging treatment, helps to reset the body’s hormonal clock and can reverse or delay the effects of aging and thus keeping young.25.5.3 The Genetic Control Theory
This theory states that animals are born with a unique genetic code, a predetermined tendency to certain types of physical and mental functioning, and that the genetic inheritance has a great deal to say about how quickly one becomes aged and how long lives. Each person has a biological clock. When that clock goes off, it signals the bodies first to age and then to die. However, the timing on this genetic clock is subject to enormous variation, depending on what happens with the growth and on how one actually lives (quality of life, feeding, sanitation and health care practices). This theory should be restricted to cases where there is a specific control of the onset of aging by an identifiable metabolic process and a functional role for senescence can be demonstrated for that species, e.g. the rapid aging and death of the Pacific salmon after spawning (Figure 25.3).
25.5.4 Waste (Undegradable By-products of Metabolism) Accumulation Theory of Aging
In the course of the lifespan, cells produce more waste than they can properly eliminate. This waste can include various toxins (free radicals, histones, aldehydes, lipofuscins), which, when accumulated to a certain level, can interfere with normal cell function, ultimately killing the cell.
The only mechanism by which these agents are diluted in the cells is cell division. This only applies to replicative cells. The challenge for multicellular organisms, such as the animal is that many cell types lose replicative capacity or divide slowly, even though they remain active throughout the lifespan. These cells, including cardiomyocytes and brain neurons, accumulate metabolic waste that eventually affects normal cell functioning. A common by-product of cellular metabolism seems to be lipofuscin (LF). Lipofuscin (age pigment) is a brown-yellow, electron-dense, autofluorescent material that accumulates progressively over time. LF is composed of highly oxidized cross-linked macromolecules (proteins, lipids, and sugars) with multiple metabolic origins. LF cannot be digested in the ubiquitin- proteasome system because of highly oxidized proteins and polymeric and highly cross-linked nature. LF cannot be cleared by exocytosis and accumulates within the lysosomes and cell cytoplasm of long-lived post-mitotic and senescent animal cells. LF inclusions from different animal and human tissues (myocardium, brain, liver, thyroid) are composed mainly of proteins and lipids, 30-70% and 20-50%, respectively. A small amount of carbohydrates (4.7%) and traces of metals are also found in LF granules. The accumulation of non-degradable material can occur in the intra- and extracellular environments. Among the extracellular deposits found in humans, cholesterol-containing plaques and their oxidized derivatives in blood vessels, as well as protein polymers, such as β-amyloid in the central nervous system.In unstressed situations, protein homeostasis is balanced by folding and stabilization of proteins by chaperones of the HSP family and the controlled degradation of proteins by the proteasomal system. The proteasome exists in different forms, and its activity is modulated by multiple regulators. The 20S core proteasome contains the proteolytic activity and selectively degrades a multitude of oxidized proteins, as well as other substrates, in an ubiquitin- and ATP-independent manner.
When the core 20S proteasome combines with two 19S regulators, the 26S proteasome is formed, which selectively removes polyubiquitinated proteins. Under stress conditions, and therefore most notably during aging, the balance between protein damage and clearance of damaged proteins is disturbed, leading to a malfunctioning of proteostasis and an accumulating mass of oxidized proteins, aggregate and aggresome formation, and finally to the accumulation of highly cross-linked materials such as lipofuscin, compromising cell viability. Accumulation of aggregates in post-mitotic cells seems to be especially dramatic since they are not able to dilute this material by cell division.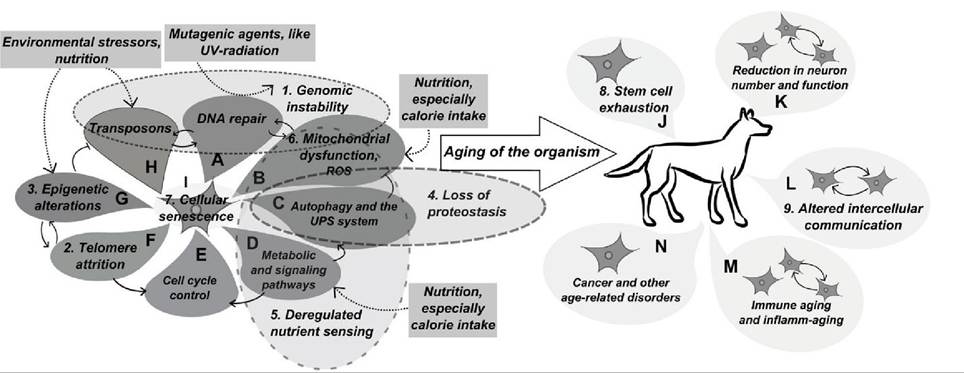
FIGURE 25.3 Main mechanisms of aging.
Consequences of oxidative stress on proteins, the pro- teasomal system, and transcription factors and formation of age pigment lipofuscin. When proteolytic capacity declines below a critical threshold of activity required to cope with oxidative stress, the final consequence is the accumulation of aggregated proteins which may, instead, cross-link with one another or form extensive hydrophobic bonds. This material might undergo further reactions and finally form the age pigment lipofuscin, which has toxic properties and accumulates in the lysosomal system. It is likely that spontaneous modification of proteins also plays a role in the formation of metabolic debris. Metabolic waste also includes, to a certain extent, spontaneously modified sugar-bound proteins, mainly glucose molecules. Glycation involves interaction between the amino groups of lysine and the aldehyde groups of glucose via a Schiff base reaction. It is followed by rearrangement of the double C=N bond, known as Amadori products, to yield a wide range of advanced glycation end-products such as glucosepane. The main consequence of spontaneous glycation is impaired elasticity, which is essential to blood vessels.
In addition, spontaneous glycation affects protein functioning. This process well describes the concept of accumulation of metabolic waste that promotes aging.25.5.3 Errors and Repairs Theory
In 1963, Dr. Leslie Orgel, a British Chemist, suggested that “an error in the machinery for making protein could be catastrophic.” The production of proteins and the reproduction of DNA sometimes are not carried out with accuracy. The body’s DNA is so vital that the natural repair processes kick in when an error is made. But the system is incapable of making perfect repairs on these molecules every time, and the accumulation of these flawed molecules can cause diseases and other age changes to occur.
25.5.4 Autoimmune Theory of Aging
(Immunosenescence)
This theory was proposed by an American scientist, Dr. Roy Walford in the year 1969. Aging is associated with declines in adaptive and innate immunity established as immunosenescence. With age the system’s ability to produce necessary antibodies that fight disease declines, as does its ability to distinguish between the antibodies and proteins. In a sense, the immune system becomes selfdestructive and reacts against itself. Thus, elderly individuals usually present chronic low-level inflammation, higher infection rates and chronic diseases. These theories suggest that with age comes decreased ability to recognize “self” from “nonself”. Antibody structure becomes modified with aging, causing them to acquire antigenic potential against normal body cells. Cells hidden during embryonic development appear later in life. Ab-Ag complexes from previous normal responses cause cumulative lesions characteristic of aging. Medications may form complexes with body proteins that are identified as foreign.
25.5.5 Cross-Linkage Theory of Aging
Developmental aging and cross-linking were first proposed in 1942 by Johan Bjorksten, US Chemist. He applied this theory to aging diseases such as sclerosis, a declining immune system and the loss of elasticity in the skin.
The cross-linking hypothesis is based on the observation that with ageproteins, DNA, and other structural molecules develop inappropriate attachments or cross-links to one another. These unnecessary links or bonds decrease the mobility or elasticity of proteins and other molecules. Denatured proteins are irreversibly altered. Denaturation is caused by cross-links between peptide strands within a protein or between proteins. With age comes new crosslinks, leading to irreversible protein structural changes and altered protein functioning.It is thought that these cross-links begin to obstruct the passage of nutrients and waste between cells. Proteins that are damaged or are no longer needed are normally broken down by enzymes called proteases. However, the presence of cross-linkages inhibits the activity of proteases. These damaged and unneeded proteins, therefore, stick around and can cause problems. One of the main ways crosslinking occurs is through a process called glycation. Glucose molecules can stick to proteins, then transform into brownish molecules called advanced glycation end products (AGEs) When both of the sticky ends of AGEs adhere to neighboring proteins, they form permanent cross-links that disable the proteins’ functions. Commonly affected body proteins a. Enzymes b. Collagen (fibrosis) c. Elastin d. Ground substance.
Cellular dysfunctions in aging or in age-related diseases by oxidative stress imbalance. 1. Cell metabolism generates reactive oxygen species (ROS) and reactive nitrogen species (RNS), which in turn causes oxidative/nitrosative damage. 2. Proteins are the most affected macromolecules by oxidative stress, undergoing several modifications that avoid their being correctly degraded and recycled by the proteasome, thus generating impaired protein function. 3. Oxidative stress also directly affects cytoskeletal proteins, causing structural damage and signaling alterations. 4. On affecting the mitochondria, oxidative stress alters energy production and 5. on affecting peroxisomes, oxidative stress alters correct metabolic functioning. 6. Oxidative stress also affects the cellular membrane. 7. Finally, all of the previously mentioned affections cause an alteration in the transcriptional activity of the cell, leading to an altered gene expression that in turn leads the cell to the aging process or to degenerative disease.
25.5.8 The Mitochondrial Free
Radical Theory of Aging
With aging, mitochondria become highly susceptible to morphological changes. These changes result in reduced function due to oxygen radical damage, which eventually causes the aging of the organism (Fakouri et al., 2019). Sites of ROS production at the mitochondrial electron transport chain. The figure shows the four complexes of the respiratory chain (CxI-IV). ROS are mainly produced at CxI and CxIII. Rotenone (ROT) blocks electron transport from ubiquinone (Q) to complex I, thus avoiding complex I ROS production when succinate is used as substrate. AA, anti- mycin A. The Mitochondrial Free Radical Theory of Aging The mitochondrial respiratory chain, illustrating electron transfer from NADH and FADH2 to oxygen. The primary function of mitochondria is respiration, which promotes energy production. Mitochondria break down organic compounds into water and carbon dioxide to release energy in the form of adenosine triphosphate (ATP).Mitochondrial respiration generates a proton gradient across the inner membrane and a transmembrane potential through respiratory chain complexes (I-IV), enabling electron flow from the reduction equivalents NADH and FADH2 to oxygen. Simultaneously, the energy released in the oxidation of NADH and FADH2 is used to pump H+ ions out of the matrix into the space between the outer and inner membranes. During respiration, oxygen is reduced in several stages, producing a superoxide radical and hydrogen peroxide. Most commonly, these molecules, known as reactive oxygen species (ROS), remain bound to cytochrome c oxidase until the reduction of oxygen to water is completed. In contrast to the common sequence of oxygen reduction by cytochrome c oxidase, oxygen molecules can occasionally form superoxide species by reacting with the reduced components of the electron transport chain. This typically occurs at the level of complexes I and III in the respiratory chain (Figure 25.4).
25.6