Passive Transport of K+ Across the Plasma Membrane Creates an Electrical Potential
As just discussed, gated ion channels can convert chemical information into electrical information. Electrical signaling in the animal body is the result of electrical imbalances maintained across the plasma membrane of virtually all cells: cells maintain an electrical potential difference across their plasma membrane.
That is, the cell membrane is a battery; if one attaches electrodes to the two ends of a battery, or to the inside and outside of a cell, one finds a voltage difference between the two ends or sides. If one provides a path for electrical charges to move—a metal wire containing free electrons in the case of a battery, or a membrane channel through which ions can move in the case of the cell—an electrical current flows from higher to lower electrical potential. The diversity of battery-powered devices in our society suggests how many ways this electrical potential can be exploited. The physiology of animals also exploits the baseline electrical potential across the plasma membrane, called the resting membrane potential. The word “resting” is added to distinguish the baseline potential from the instantaneous values of membrane potential during the passage of membrane currents.The resting membrane potential is the indirect result of the concentration gradients of ions across the plasma membrane caused by the activity of the NaK+-ATPase. Partly, this membrane potential is a result of the asymmetry in numbers of ions pumped by the Na*,K+-ATPase. However, most of the membrane potential is caused by the passive flow of K+ through K' leak channels in response to the concentration gradient of K’ (high inside, low outside). 'Γhis concentration gradient sets up an electrical driving force (voltage) that exactly balances the concentration driving force. The concentration of K4 inside a mammalian cell is about 150 mmol; outside in the interstitial fluid, it is about 5 mmol.
As a result, K’ tends to diffuse from the cytoplasm through the leak channel to the interstitial fluid. However, when Ki alone leaves the cytoplasm without an accompanying negative ion, it causes an electrical imbalance. The exit of K, ions leaves the inside of the cell with negative charges not neutralized by positive potassium ions, and the interstitial fluid now has positive K+ ions not balanced by negative charges. The cell is building an electrical potential difference across the plasma membrane, with the cytoplasm being negative relative to the interstitial fluid.This electrical potential driving force increases until it balances the concentration driving force for K1. This situation is analogous to osmosis: the concentration-driven flow of water across a Semipermeable membrane creates a different driving force, pressure, that eventually balances the concentration driving force. Similarly, for the resting membrane potential, the concentration-driven flow of K' across the semi- permeable membrane (Semipermeable in the sense that negative ions do not accompany the K+) creates a different driving force, an electrical voltage, that eventually balances the concentration force. As in the case of osmosis, an equation is used to relate the size of the concentration gradient to the size of the electrical potential that provides an exact balance. This equation is called the Nernst equation, as follows:
Ex = RT/zF ∣n[Xoutsidβ]∕(XιnsidJ
Ex = Equilibrium potential for ion X.
RT = Gas constant ? Absolute temperature.
z = Electrical valence for the ion, +1 for Na* and K∖ -1 for CΓ, and so forth.
F = Faraday constant = number of coulombs of electrical charge in a mole of ions = 96,500 coulombs/mol.
hi = Natural logarithm (i.e., log to base e).
(XJ = Concentration of ion X.
A simpler form of this equation can be written by taking advantage of the fact that R and F are constants, T is almost constant under physiological conditions, and the natural log of a number is 2.3 times the common log (log∣0), as follows (mV, millivolts):
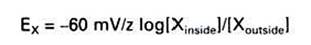
Because the state of balance between the electrical driving force and the concentration driving force is equilibrium, the value of the electrical potential is called the equilibrium potential of the ion. Given the concentrations above for K+ inside (150 mmol) and outside (5 mmol) the cell, the equilibrium potential for K’ is:

Indeed, the measured resting membrane potential across a human muscle cell is -90 mV.
Several aspects of this important equation are worth discussing. If the equilibrium potential for a particular ion is the same as the measured membrane potential, the net driving force for the ion is zero. In this case, there is no net movement, even in the presence of wide-open channels, to provide a path through the membrane. However, for any gradient of a specific ion, if the measured membrane potential is not the equilibrium potential of that ion, there is a driving force for the transport of that ion. That is, when the membrane potential is anything other than the equilibrium potential, that ion will flow across the membrane if an appropriate channel is open. Thus the equlibrium potential for an ion provides a “baseline” for comparison with the actual membrane potential to determine whether an ion will tend to move across the plasma membrane. If the measured membrane potential has the same sign but is larger in magnitude than the equilibrium potential, the ion flows in the direction of the electrical potential. If the sign is the same but the magnitude lower, the concentration driving force determines the direction of flow of the ion. If the measured potential has the opposite sign of the equilibrium potential, both electrical and concentration forces arc acting on the ion in the same direction. Flow of ions across the plasma membrane (i.e., electrical current) in response to the balance of force between concentration and voltage produces the electrical changes in neurons that underlie the nervous system, as discussed in Chapter 4.
It would be reasonable but incorrect to assume that the transport of ions required to set up the electrical potential measurably alters the concentration gradient. This is untrue because of the large amount of energy required to separate electrical charges. The separation of charge arising from the transport of a few ions balances the energy of quite substantial concentration gradients. Indeed, so few ions move that they cannot be measured by chemical means.
Thus, electrical, not chemical, measurements are used routinely to assess transport of ions in cells. The measurable voltage changes caused by immeasurably small concentration changes of ions means also that the electrical phenomena at the membrane persist for many hours, even if the Na’,K*-ATPase is inactivated by a toxin. That is, an existing concentration gradient of K+ would require hours to dissipate at the rate of K* leakage characteristic of the plasma membrane. Using the membrane battery analogy, the Na11K+-ATPase is a battery recharger. A portable radio does not require the minute-to-minute services of a battery recharger. Enough energy is stored in the battery to operate the radio for an appreciable period, although the battery recharger is needed eventually. Similarly, enough energy is stored in the K+ concentration gradient to maintain the membrane potential for a period of time. The Na11K+-ATPase is not required on a minute-to-minute basis, although it is needed ultimately to maintain the concentration gradient on which the resting membrane potential depends.