PATTERNS OF ARTERIAL DISTRIBUTION
We have already mentioned certain more obvious features of arterial distribution: the increase in total crosssectional area at each branching, the variation in the angle of branching, the preference for protected courses
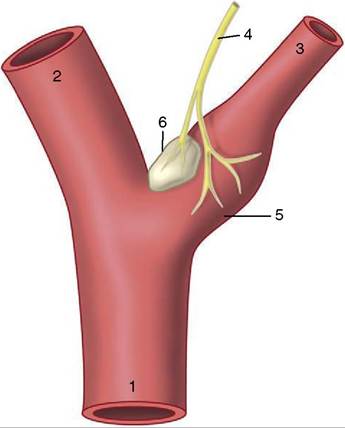
Figure 7-32 Baroreceptors and chemoreceptors at the origin of the internal carotid artery.
1, Common carotid artery; 2, external carotid artery; 3, internal carotid artery; 4, carotid sinus branch of the glossopharyngeal nerve; 5, carotid sinus (baroreceptor); 6, carotid body (chemoreceptor).within the limbs, and the generosity of interarterial anastomoses (p. 27). Amplification of the description of certain features is required.
Collateral Circulation
Few arteries of any size proceed to their terminations in capillary beds without first detaching side or collateral branches. Most collateral branches, whether large or small, connect with their neighbors, although the profusion of anastomoses may not be apparent on dissection because so many are concealed within muscles and other organs (Figure 7-33). The anastomoses enlarge when the bloodstream is diverted from its normal route by occlusion of a principal trunk; initially the widening is due to relaxation and stretching of the wall but later is due to reconstruction of the anastomotic links. Thus, provided that sufficient blood can pass in the meantime, tissues deprived of their usual sources of supply generally survive, though possible temporary loss of function of the ischemic parts may occur. Experiments have shown that in healthy dogs even the aorta can be ligated (caudal to the origin of the renal arteries) leaving a fair, perhaps 50%, expectation of survival. This does not mean that any artery can be ligated with impunity. The ability to develop an adequate collateral circulation is increased when the obstruction develops slowly; it is lessened by sudden onset, aging, or frankly pathological changes in the vessel wall.
Some arteries have a patency that is essential: interruption to flow produces an infarct, the death of a block of tissue (typically shaped like a cone about the vascular axis). These arteries, known as end-arteries, are paradoxically more numerous among smaller arteries than their parent trunks, which generally have more extensive collateral connections. By strict definition, the endartery is a rarity, but “functional” end-arteries, in which the collateral connections are of insufficient caliber, are more common (Figure 7-34). It is impossible to assess the adequacy of collateral circulation from purely morphological evidence; for example, although intramuscular arteries appear to anastomose freely, occlusion of one frequently leads to local necrosis. Other good examples of arteries in which anastomoses are poor are the central artery of the retina and many small vessels
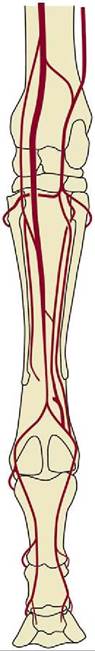
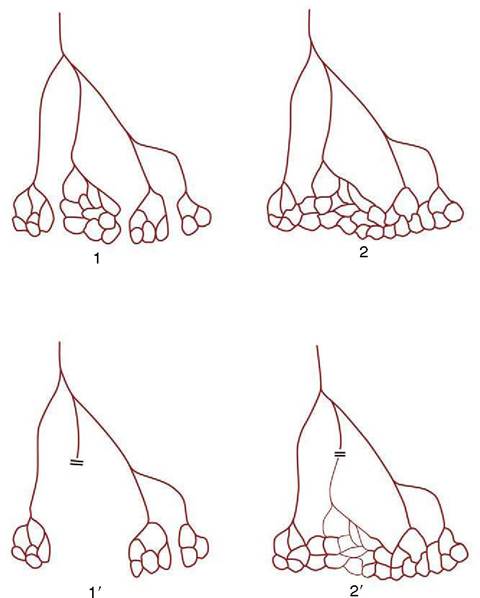
Figure 7-34 True (1) and functional (2) end-arteries. Closure of an end-artery leads to necrosis of the tissue it supplies (1 ). In the case of a functional end-artery (2), a potential but inadequate alternative route exists (2').
Figure 7-33 This illustration of the arterial pattern of the equine limb shows the generosity of interarterial anastomoses.
within the brain; the consequence of their obstruction may be immediate and catastrophic: destruction of the retina or the death of a nucleus or tract along with permanent sensory or motor disability. This may be contrasted with the freedom of anastomoses between the major arteries that conjoin to form the arterial circle on the ventral surface of the brain. Anastomoses between finer branches of the coronary arteries are also poor and usually incapable of maintaining an adequate collateral circulation; even so, not all coronary embolisms are fatal.
Much may depend on the size and specific site of the infarct and on immediate medical care.Anastomoses between small arteries within the limbs are especially numerous in the regions of the joints and sometimes form visible networks or retia; a prominent example exists over the dorsal aspect of the carpus of the horse (rete carpi dorsale).
The retia just described are not to be confused with the so-called retia mirabilia of more restricted occurrence. Retia mirabilia are found where a main trunk splits more or less at once into a leash of parallel vessels. In one variety the parallel trunks later reunite; this is a “bipolar” arrangement found on the arteries to the brain (in certain species) (Figure 7—35/7) and, on a diminished scale, in the renal glomeruli (see Figure 5-28). Other examples are “unipolar,” that is, the branches remain separate. Examples are found within the limbs of slow-moving arboreal creatures (sloths, lemurs) and in the thoracic cavity of whales and other diving mammals. No convincing explanations exist of the adaptive value of most of these; the renal glomeruli, however, are the obvious exception (p. 181).
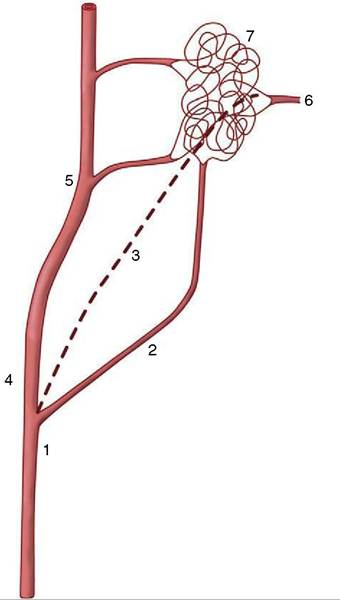
Figure 7-35 A rete mirabile interposed on the blood supply to the bovine brain. 1, Common carotid artery; 2, occipital artery; 3, internal carotid artery (regresses after birth); 4, external carotid artery; 5, maxillary artery; 6, branch from rete to arterial circle of the brain; 7, rostral epidural rete mirabile.