Physiology of Respiration
Ventilation
Ventilation is the process by which air is moved into (inspiration) and out of (expiration) the lungs. Alveolar ventilation is a more specific term that refers to the movement of air into and out of alveoli.
Alveoli are the primary site of gas exchange in the lungs, so alveolar ventilation is the critical component of this function.Panting is a mechanism to dissipate heat. Panting is characterized by an increased ventilatory rate but with a reduced tidal volume (volume of air moved during each breath). Alveolar ventilation increases minimally in panting animals, because the increase in air movement is primarily in the upper airways that are not sites of gas exchange. These airways are considered to be anatomic dead space. Physiologic dead space includes the anatomic dead space and any alveoli in which normal gas exchanges cannot occur.
As described earlier, there is no physical connection between the visceral and parietal pleural surfaces (except at the hilus), and the closed pleural cavity between them is a potential space filled with a small amount of fluid. The hydrostatic pressure in the pleural cavity is always slightly negative relative to atmospheric pressure (Fig. 19-10), and this small negative pressure exerts a pulling force to keep the lungs expanded. Enlargement of the thoracic cavity during inspiration reduces the already negative pressure in the pleural cavity, which further expands the lungs (Fig. 19-10).
During inspiration, air moves in through the upper airways and down to the alveoli because of a hydrostatic pressure gradient (i.e., atmospheric pressure) between the outside air and the air passages in the lungs (intrapulmonic pressure). This gradient is generated by lung expansion. Expansion increases the volume of
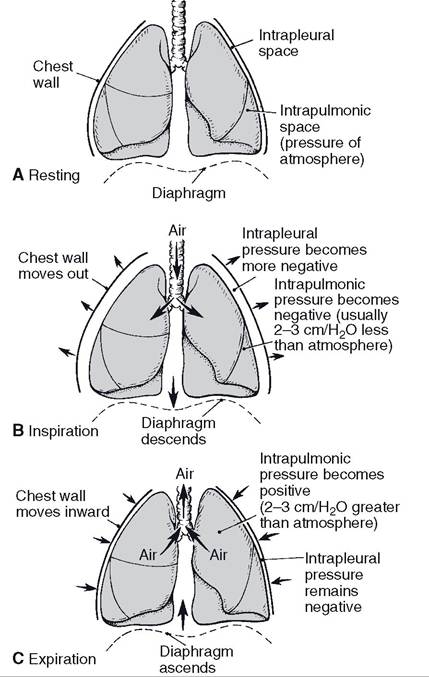
Figure 19-10.
Phases of ventilation. A) No air movement (resting). B) Air moves from the atmosphere to the intrapulmonic space (inspiration). C) Air moves from the intrapulmonic space to the atmosphere (expiration). (Reprinted with permission from Bullock B.L. Pathophysiology. 4th ed. Philadelphia: Lippincott, 1996.)the air passages in the lungs but reduces the pressure in them. The inverse relationship between volume and pressure of a gas is expressed as Boyle’s law. Figure 19-10 illustrates the changes in intrapulmonic and intrapleural pressures during inspiration and expiration. The negative (inspiration) and positive (expiration) values for intrapulmonic pressures are relative to atmospheric pressure (760 mm Hg at sea level), so negative pressure is less than atmospheric and positive is greater than atmospheric.
Enlargement of the thoracic cavity is accomplished by contraction and flattening of the dome-shaped diaphragm and a forward and outward movement of the ribcage by the contraction of appropriate thoracic muscles. These are skeletal muscles innervated by somatic motor nerves. After inspiration, the pressure in the pleural cavity remains at its lowest point until expiration begins and the thoracic cavity begins to return to its original volume.
Expiration in a resting animal is a passive process that does not require muscle contraction. Relaxation of the muscles contracted during inspiration permits the intrinsic elastic properties of the lungs and the thoracic wall to recoil to their original volume. The return to the original volume increases the intrapulmonic pressure so that it is greater than atmospheric pressure, and air is forced out of the lungs.
Forced expiration is an active process that forces more air from the lungs than would occur during a normal passive expiration. Forced expiration requires contraction of abdominal muscles to force viscera against the diaphragm and contraction of other muscles to pull the ribs caudad. Both of these actions reduce the size of the thoracic cavity and permit recoil of the lungs to a smaller volume than typical for resting expiration.
This causes a further increase in intrapulmonic pressure and forces more air out than would occur with passive expiration. Even with the most forceful expiration, a residual volume of air still remains in the lungs. Vital capacity is the maximal amount of air that can be expired after a maximal inspiration.Both the upper airways and the air passages in the lungs resist airflow through them. Similar to a blood vessel, the resistance of an air passage to airflow is inversely proportional to its diameter. Most of the air passages in the lungs have smooth muscle in the walls, and the constriction or relaxation of this smooth muscle determines the resistance to airflow. Much of this smooth muscle has β2-adrenergic receptors that produce smooth muscle relaxation when stimulated. Sympathetic stimulation during exercise reduces resistance to airflow and promotes alveolar ventilation.
Airway smooth muscle constricts in response to stimulation by histamine or leukotrienes. These substances are released during allergic reactions from mast cells around airways. The resulting increase in airway resistance makes it more difficult to move air, and a more forceful skeletal muscle contraction is needed to move a given volume of air. This greatly increases the work of breathing. Heaves is a condition in horses that is characterized by labored or difficult breathing, and one potential cause is chronic exposure to environmental allergens, which results in chronic obstructive airway disease. The chronic need for increased skeletal muscle force may induce visible hypertrophy of the abdominal muscles (heave line).
Surface tension, a property of fluids, results from the cohesive forces that tend to pull or hold the molecules of a fluid together. An example is the tendency of a layer of fluid on a flat surface to come together and form a bubble rather than remaining as a thin, flat layer. A thin layer of fluid lines the microscopic alveoli. The surface tension of this fluid tends to draw the walls of alveoli together and collapse them.
The alveolar fluid contains the pulmonary surfactant, a combination of substances that reduces surface tension. The reduction in surface tension promotes alveolar stability and makes alveolar expansion during inspiration easier. Production of pulmonary surfactant does not begin until late in gestation. Neonates born prematurely may have insufficient amounts of surfactant, which results in labored breathing.in normal animals a soft rustling sound associated with air movements may be heard with a stethoscope. Abnormal lungs, for example lungs with abnormal amounts of fluid in the airways, may produce exaggerated sounds termed rales. Pleuritis (pleurisy), or inflammation of the pleura, may produce a rasping sound because of the roughened surfaces rubbing together. Hydrothorax (fluid in the pleural cavity) may also result from pleuritis.
Gas Exchange
The oxygen and carbon dioxide concentrations in air can be described in two ways: partial pressures and percentages. Room air is approximately 21% oxygen and 0.3% carbon dioxide. The primary component of room air is the inert gas nitrogen (about 78%). The partial pressure of an individual gas in a mixture of gases is the product of the percentage of the individual gas in the mixture and the total barometric or atmospheric pressure. Thus, at sea level, where atmospheric pressure is 760 mm Hg, the partial pressure of oxygen in room air is approximately 160 mm Hg.
The partial pressure of an individual gas in a mixture is one factor that determines the amount of the gas that will dissolve in a liquid (such as blood plasma). The partial pressure of a gas in a mixture can be viewed as the driving force that moves molecules of an individual gas from the air into the liquid when the liquid is exposed to a gas mixture. Thus, because partial pressures depend on both total atmospheric pressure and the percentages of individual gases, both of these factors also determine the amount of an individual gas that can be dissolved in a liquid.
The unit of measurement of the amount of a gas dissolved in a liquid is in millimeters of mercury.Gas Exchange in the Lungs. Gas exchange between the blood and alveolar air in the lungs occurs across the walls of alveoli. At its thinnest point, the alveolar wall barrier between blood plasma and alveolar air consists of the endothelial cell of pulmonary capillaries, a type i squamous epithelial cell lining the alveoli, and a fused basement membrane contributed by both cells (Fig. 19-11). Gases readily move back and forth across this very thin and fragile structure. Any abnormality that thickens this barrier (e.g., pulmonary edema with an accumulation of extracellular fluid in the alveolar wall) can greatly reduce the efficiency of exchange. Oxygen exchange is usually affected first because its solubility is much less than that of carbon dioxide.
Exchange begins as soon as blood enters a pulmonary capillary from pulmonary arterial vessels and continues until equilibrium between alveolar air and plasma is reached. Figure 1912 shows typical values for partial pressures of
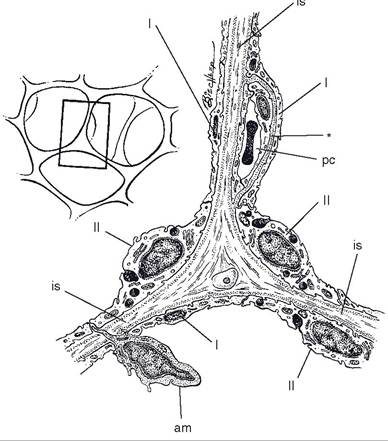
Figure 19-11. Schematic representation of three adjacent alveoli. I- type I alveolar epithelial cell; II- type II alveolar epithelial cell; is- interalveolar septum; pc- pulmonary capillary; am- alveolar macrophage. Note the merger of the basal laminae of the pulmonary capillary and alveolus (*). (Reprinted with permission of Wiley-Blackwell from Dellmann H.D. Textbook of Veterinary Histology. 4th ed. Philadelphia: Lea & Febiger, 1993.)
oxygen and carbon dioxide in alveolar air and a pulmonary capillary. Plasma entering pulmonary capillaries from the pulmonary arteries contains the highest concentration of carbon dioxide and the lowest of oxygen. Because of the continuous gas exchange in the pulmonary capillaries, alveolar air contains less oxygen and more carbon dioxide than inspired air (Po2 of 160 mm Hg and PCO2 of 0.23 mm Hg for inspired air).
To be most efficient, the rate of pulmonary artery blood flow into an area of the lung must be balanced with the rate of air movement in and out of the alveoli in the same area. To appreciate the importance of this balancing, consider an extreme case in which one lung is collapsed so that air movement is impossible, but the collapsed lung receives the same amount of blood flow as the inflated normal lung. The lack of airflow in the collapsed lung means that
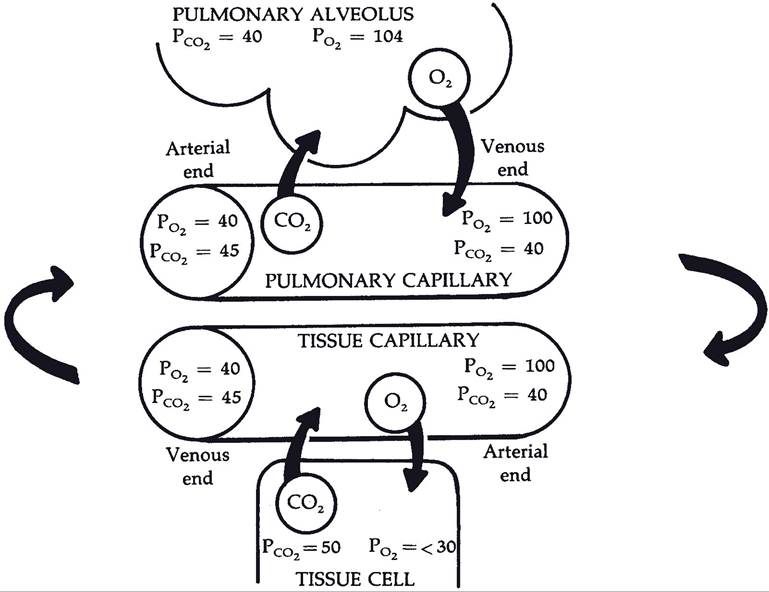
Figure 19-12. Direction of diffusion of oxygen (O2) and carbon dioxide (CO2), as shown by arrows. Values for gases in alveolar air and in fluids (plasma and intracellular fluid) are expressed in millimeters of mercury. Note changes in values as blood flows from arterial to venous end of the two types of capillaries. (Reprinted with permission of Wiley-Blackwell from Reece W.R. Physiology of Domestic Animals. 2nd ed. Baltimore: Williams & Wilkins, 1997.)
any alveolar air is stagnant and contains high levels of carbon dioxide and low levels of oxygen. Gas exchange in the collapsed lung does not occur, and levels of oxygen and carbon dioxide are the same as those entering. The blood exiting the collapsed lung mixes with an equal volume of blood from the intact lung, so that blood reentering the heart from the pulmonary circulation is deficient in oxygen and has an excess of carbon dioxide.
Within the lungs, a unique type of vascular mechanism functions at the level of small arterial blood vessels to balance blood flow and airflow. This mechanism, local hypoxic (low oxygen) vasoconstriction, produces local vasoconstriction in response to low levels of alveolar oxygen (such as with poor alveolar ventilation). The vasoconstriction reduces blood flow into the area of poor ventilation and shunts blood into better-ventilated areas of the lungs. it is not clear how low levels of alveolar oxygen are detected and what vasoactive agent or agents are responsible for the vasoconstriction.
The hypoxic vasoconstriction mechanism operates well on a local basis to redirect blood flow into different areas of the lungs. However, when both lungs are exposed to low oxygen levels, such as at high altitudes, the mechanism produces a general increase in vascular resistance throughout both lungs. Pulmonary hypertension (high pulmonary circulation blood pressure) results, and the right side of the heart must work harder to pump blood through the lungs. Right heart failure with peripheral edema can result if the right heart cannot compensate for the increased resistance. A syndrome with pulmonary hypertension, right heart failure, and peripheral edema resulting from exposure to high altitude is recognized in cattle as brisket disease, in which edema collects in the pendulous brisket.
Gas Exchange in the Tissues. Cells in peripheral tissues consume oxygen and produce carbon dioxide during normal metabolism. This maintains relatively low oxygen and high carbon dioxide concentrations (partial pressures) in the extracellular fluid around capillaries. As arterial blood enters capillaries, partial pressure gradients promote the diffusion of oxygen out of the blood to the interstitial fluid and carbon dioxide from the interstitial fluid into the blood (Fig. 19-12).
Gas Transport in Blood
Both oxygen and carbon dioxide dissolve in plasma, and the partial pressures of each are a measure of the amount dissolved. However, the quantity of each gas that is transported as a dissolved gas is very small compared with the amounts of each transported in other forms in the blood. Only 1.5% of the total oxygen and 7% of the carbon dioxide are dissolved.
Most oxygen in the blood (98.5%) is found chemically bound to hemoglobin in erythrocytes (see Chapter 15). Figure 19-13A, illustrates the relation between the partial pressure of oxygen and the percent saturation of hemoglobin by oxygen. At partial pressures of oxygen normally found in alveolar air (about 100 mm Hg), hemoglobin is almost completely saturated with oxygen (i.e., hemoglobin molecules cannot bind any additional oxygen). Normally, the hemoglobin in erythrocytes is almost completely saturated with oxygen as blood passes through the pulmonary capillaries. At partial pressures of oxygen typically found in venous blood (40 mm Hg) a great deal of hemoglobin still has oxygen bound to it (Fig. 19-13A).
several factors affect the ability of hemoglobin to bind chemically with oxygen. An increase in temperature, a reduction in pH, or an increase in the concentration of carbon dioxide reduces the ability of hemoglobin to bind oxygen. These factors alter the relation between hemoglobin saturation and the partial pressure of oxygen so that the saturation is less for any given partial pressure. Figure 19-13B illustrates the effect of
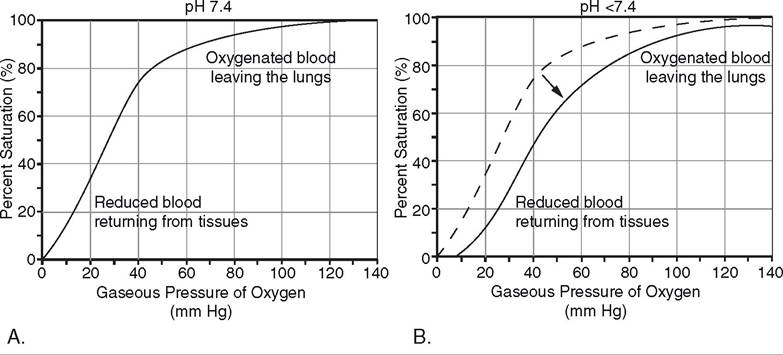
Figure 19-13. A) An oxygen-hemoglobin dissociation curve that illustrates the percent saturation of hemoglobin by oxygen at different partial pressures of oxygen. B) Effect of lowering the pH from 7.4 to 7.2 on the oxygenhemoglobin dissociation curve.
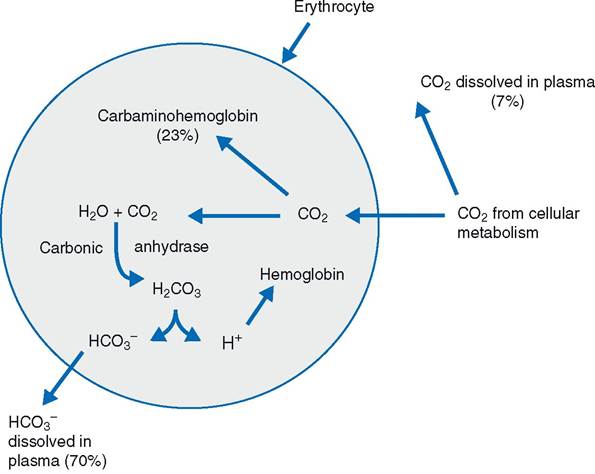
Figure 19-14. Gas exchange in systemic tissues and transport of carbon dioxide in the blood. Percentages reflect the different modes by which carbon dioxide is transported from systemic tissues to the lungs.
lowering pH on hemoglobin saturation, and the effects of increases in temperature or carbon dioxide are similar. High temperature, low pH, and high carbon dioxide occur in tissues with high metabolic rates (e.g., exercising skeletal muscle). The effects of these factors on the relation between hemoglobin and oxygen is useful, because more oxygen is liberated from hemoglobin and delivered to the metabolizing cells when blood passes through such areas.
Almost all of the carbon dioxide (93%) that enters the blood in the systemic circulation diffuses into erythrocytes (Fig. 19-14). Some of the carbon dioxide (23%) chemically combines with the hemoglobin in the erythrocytes to form Carbaminohemoglobin. When the erythrocytes carrying the carbaminohemoglobin reach the pulmonary capillaries in the lungs, the reaction is reversed so that the carbon dioxide can diffuse into alveoli to be expired. Most (70%) of the carbon dioxide that enters the erythrocytes is converted to carbonic acid under the influence of the enzyme carbonic anhydrase. The carbonic acid rapidly dissociates into a hydrogen ion and a bicarbonate ion in the erythrocyte. The hydrogen ion is buffered by hemoglobin in the erythrocyte, and the bicarbonate ion leaves the erythrocyte and enters the plasma. it is in this form (bicarbonate ion in plasma) that most carbon dioxide is transported from the peripheral tissues by the blood to the lungs (Fig. 19-14). Within the lungs, the reactions are reversed so that carbon dioxide can be reformed and be expired from the alveoli.
Control of Ventilation
Contraction and relaxation of skeletal muscle generates the forces to move air in and out of the lungs. Although the skeletal muscles of respiration can be consciously controlled, as illustrated by voluntarily holding the breath, normal ventilation is almost entirely reflexive.
The respiratory reflex centers consist mainly of three bilateral groups of nerve cells in the brainstem that have a definite effect on respiration when stimulated electrically. one of these, the medullary rhythmicity area, is responsible for setting respiratory rate, and it consists mainly of an inspiratory center. Neurons in this center are tonically active, firing at an inherent rhythmic rate by regular variations of their membrane potentials. Expiratory neurons are also in this area; however, they do not discharge spontaneously and so are normally active only during a forced expiration.
stimulation of the inspiratory center leads to contractions of the diaphragmatic and intercostal muscles via neural connections through the spinal cord and phrenic and intercostal nerves, respectively. Feedback circuits between the inspiratory center and the other two neural centers relax these muscles and allow for passive expiration. The interplay among these three centers provides for regular intermittent rhythmic breathing at rates appropriate for each species during eupnea (normal quiet breathing).
The tonic activity of the inspiratory center is regulated by neural input from a variety of sites. in resting animals, the most important neural input is from central chemoreceptors in the medulla of the brainstem. These receptors respond to hydrogen ion concentration changes in the interstitial fluid of the brain and stimulate the inspiratory center to increase ventilation when the hydrogen ion concentration increases. Because carbon dioxide from the blood readily diffuses into the interstitial fluids of the brain, and in body fluids is in equilibrium with carbonic acid, an increase in blood carbon dioxide increases the hydrogen ion concentration in the brain and stimulates ventilation. The effects of changes in blood carbon dioxide on ventilation are so pronounced that blood carbon dioxide is considered to be the most important regulator of ventilation in most conditions.
Animals sometimes hyperventilate (breathe abnormally rapidly) during the induction of general anesthesia because they are excited. As a result of the hyperventilation, their blood carbon dioxide may be significantly reduced. The low blood carbon dioxide concentration means that the primary stimulus for normal ventilation is lost. These animals may undergo a period of apnea (cessation of breathing) until blood carbon dioxide levels are restored by metabolism.
Another group of chemoreceptors, the carotid and aortic bodies, also provide neural input to the inspiratory center. These peripheral chemoreceptors detect changes in arterial blood hydrogen ion concentration and oxygen content. increases in hydrogen ion concentration or reductions in blood O2 content initiate neural inputs to increase ventilation. However, the effects of these peripheral chemoreceptors are less pronounced than those of the central chemoreceptors, so changes in blood hydrogen ion concentration or oxygen content must be severe to override the effect of blood carbon dioxide.
Further regulating the inspiratory center and breathing rhythmicity is a reflex arc involving stretch receptors in the lung parenchyma, visceral pleura, and bronchioles. These receptors are stimulated as the lung inflates during inspiration, and afferent impulses are transmitted up the vagus nerves into the brainstem, where the inspiratory center is inhibited. This is the Hering-Breuer reflex, which reinforces the action of the other center to limit inspiration and prevent overdistension of the lungs.