PREGNANCY
1. Know the length of gestation for each of the domestic species (see Table 15-1).
2. What is a sperm reservoir? Where are important ones located?
3. What is capacitation? Name one capacitation change.
4. What is the zona reaction associated with fertilization? Where does fertilization normally occur?
5. What is uterine milk?
6. What is implied by implantation?
7. What is placentation? What membranes compose the fetal placenta?
8. Know the relationship of the placental membranes to each other and to the fetus and mother. Where are the branches of the umbilical arteries and veins located?
9. What is a persistent urachus?
0. Which animals have a cotyledonary placenta? What composes a placentome?
11. Which steroid hormone predominates during pregnancy? Where is it produced? Do the sources and duration of their production vary among species? When is the corpus luteum source needed by all species?
2. What function is served by pregnant mare serum gonadotropin (PMSG)?
3. What are some signs of pregnancy in the cow as observed by rectal palpation?
Pregnancy is the condition of the female in which unborn young are contained within the body. Pregnancy is also called gestation, and its length is frequently known as the gestation period, extending from fertilization through birth. Its length for various domestic animals is shown in Table 15-1. Pregnancy begins with fertilization, ends with parturition, and includes the essential aspects of implantation and placentation. Before fertilization, the oocyte and sperm are transported to appropriate sites in the uterine tubes.
Transport of Oocyte and Spermatozoa
At ovulation, the fimbriae of the uterine tubes (see Figure 15-1) are in close contact with the ovaries. The contractile activity of the fimbriae directs the shed oocyte into the funnel-shaped opening of the uterine tube.
Within the uterine tube the oocyte is directed toward the uterus by cilia and by uterine tube motility.The ejaculated spermatozoa are transported to the uterine tubes by increased motility within the uterus caused by the release of oxytocin at the time of coitus and by the presence of prostaglandins in semen. The oxytocin is effective because of the uterus being primed by estrogen. Another factor that assists in transport is thought to be the presence of a negative pressure (vacuum) in the uterus. Many spermatozoa are rapidly transported to the uterine tubes after ejaculation, but it is believed that these are not the ones destined for fertilization. Their presence might be coincidental with the spread of accessory fluids throughout the tubular genitalia. The spermatozoa destined for fertilization are transported more slowly from their sites of deposition (cervical canal, uterus, vagina) to spermatozoa reservoirs. The cervix of ruminants has prominent ridges and mucosal crypts that provide an extensive secretory surface (Figure 15-20). The cervical crypts and their mucous covering aid in the physical entrapment of spermatozoa and serve as spermatozoa reservoirs. Another important spermatozoa reservoir is located at the junction of the uterine horns with the uterine tubes.
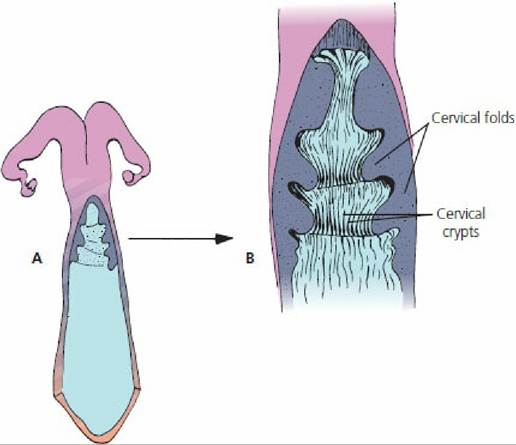
■ FIGURE 15-20 Dorsal view of the ruminant cervix. A. The cervix has been cut open and its lateral walls reflected to show the folds and crypts. B. Magnified view of the cervix. A mucous covering assists physical entrapment of spermatozoa destined for fertilization. The folds and crypts serve as sperm reservoirs and allow for capacitation of spermatozoa.
Within the spermatozoa reservoirs, the spermatozoa undergo changes necessary for later penetration of the zona pellucida and fertilization of the oocyte. These changes, known as capacitation, require several hours. One important change involves the acrosome, in which channels are established for the escape of hyaluronidase and a proteolytic enzyme; these substances are essential for penetration of the ovum.
Capacitated spermatozoa are released slowly from the spermatozoa reservoirs and proceed to the ampulla of the oviduct (dilated portion near the infundibulum) for fertilization. Ovulation occurs after the onset of estrus so that insemination is accomplished before ovulation. This allows enough capacitation time and, because the fertilizing lifespan is twice as long in spermatozoa as in oocytes, large numbers of spermatozoa are usually ready for fertilization at the time of ovulation. Oocytes retain viability for about 12 to 18 hours after ovulation in most domestic animals, and spermatozoa retain their fertilizing ability for 24 to 48 hours in the cow, ewe, and sow, for up to 90 hours in the bitch, and for 120 hours (5 days) in the mare.Fertilization
Fertilization is the fusion of male and female gametes to form one single cell, the zygote. The first step in fertilization is penetration of the zona pellucida by the spermatozoon. This involves not only the enzymes hyaluronidase and acrosin (proteolytic enzyme from acrosome), but also spermatozoon motility. Motility ceases once contact with the oocyte has been made. In most domestic species, the second maturation division (meiosis) occurs when a spermatozoon penetrates the zona pellucida, whereas the first meiosis occurred a few hours before ovulation. The zona reaction occurs after penetration of the zona pellucida and protects the oocyte from further penetration by other spermatozoa. Penetration by more than one spermatozoon (polyspermy) is deleterious to normal development of the zygote.
Pronuclei develop from the nuclei of the spermatozoon and oocyte, which is followed by fusion of respective pronuclei to form a zygote with the diploid number of chromosomes (see Chapter 1). Fertilization is complete after the fused pronuclei have disappeared and are replaced by chromosome groups united in prophase of the first mitotic division.
Zygotes usually remain in the uterine tube for 3 to 4 days before being transferred to the uterus.
Uterine motility is unfavorable for zygote survival and estrogen dominance at estrus must be changed to progesterone dominance, which occurs with the formation of the corpus luteum. Progesterone has a quieting influence on the uterus and promotes development of a glandular endometrium that can secrete uterine milk, a nutrient medium for the embryo preceding its implantation. Cell division produces a 16- to 32-cell structure known as the morula. A cavity forms within the morula by 6 to 8 days of age, and the cell mass is called a blastocyst.The period of the oocyte ends when the blastocyst attaches to the endometrium. This is the beginning of the embryonic period. The embryonic period is characterized by rapid growth; major tissues, organs, and systems develop and the major features of external body form become recognizable. The fetal period extends from this time until birth and begins at about day 45 of gestation in the cow.
Implantation and Placentation
The nutritive requirements of the developing blastocyst are satisfied by diffusion from yolk in the oocyte and by secretions of the uterine tube and uterus (uterine milk) until it becomes fixed in position in the uterus. Implantation of the embryo occurs when it becomes fixed in position and forms a physical and functional contact with the uterus. It occurs 2 to 5 weeks after fertilization. The interval is shortest for the cat (2 weeks) and longest for cattle and horses (5 weeks).
Because the embryo continues to grow, the central mass of cells becomes further removed from the surface. Diffusion of nutrients is no longer adequate and membranes develop concurrent with a circulatory system that provide for receiving nutrients from the dam. The development of extraembryonic membranes is known as placentation, and the collective name for the membranes is the fetal placenta, which consists of the chorion, allantois, and amnion. The relationship of the fetal membranes to the fetus is shown in Figure 15-21. The chorion is the outermost membrane and is the one most intimately associated with the endometrium.
The amnion envelops the fetus and contains amniotic fluid in the amniotic cavity. The amniotic fluid is derived from fetal urine from the urethra, from secretions of the respiratory tract and oral cavity, and from the maternal circulation. The amniotic fluid protects the fetus from external shock, prevents adhesion of fetal skin with amniotic membrane, and assists in dilating the cervix and lubricating the birth passage at parturition. The allantois outer layer is fused to the chorion and the inner layer of allantois is fused to the amnion. The space between the two layers of allantois is called the allantoic cavity. It is continuous with the cranial extremity of the urinary bladder by way of the urachus, which passes through the umbilical cord. When the urachus fails to close at birth, a continuous drip of urine is observed from the navel, a condition known as persistent urachus (Figure 15-22). Allantoic fluid originates from fetal urine and from secretory activity of the allantoic membrane. The fluid brings the chorioallantoic membrane into close apposition with the endometrium during early attachment and stores fetal excretory products. Branches of umbilical arteries and veins are distributed between the outer layer of allantois and the chorion.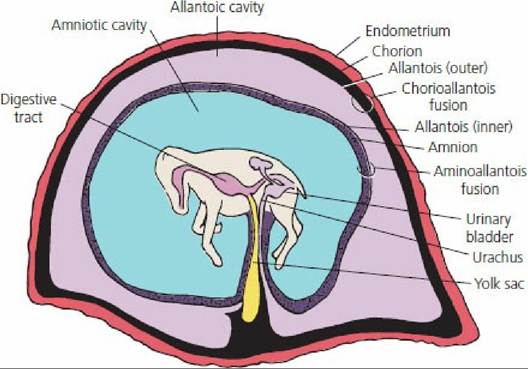
■ FIGURE 15-21 Fetus of horse within the placenta. The chorioallantois is the combination of the outer allantois with the chorion. Umbilical arteries and veins (not shown) occupy the space (blackened) between the outer allantois and chorion. The chorion is intimately associated with the endometrium. Attachment to the endometrium is not shown, and its extent varies with placental type. The inner allantois is fused with the amnion (stippled for contrast).
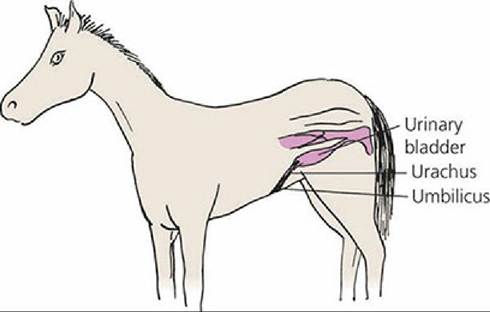
■ FIGURE 15-22 Diagrammatic view of persistent urachus in a foal.
Failure of urachus closure at birth results in a continuous drip of urine at its umbilical exit.When the attachment (extension of chorionic villi) of fetal membranes to the endometrium is continuous throughout the entire surface of the fetal membranes, it is known as a diffuse placenta. A diffuse type of placenta is found in the horse and pig (Figure 15-23A). Ruminants have a cotyledonary placenta, in which attachment occurs only at the many mushroom-like projections from the endometrium (Figure 15-23B). The fetal cotyledons are attached to the maternal caruncles, a combination known as a placentome (see Figure 15-7). The fetal placentas of the dog and cat are attached by a girdle-like band that encircles the placenta, called a zonary placenta (Figure 15-23C). The human placenta attachment is confined to a disk-shaped area and is called a discoidal placenta (Figure 15-23D).
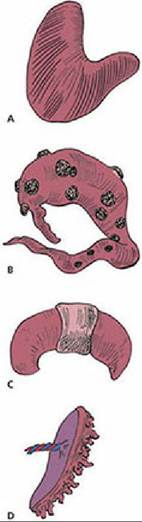
■ FIGURE 15-23 Placental types according to the distribution of chorionic projections (villi) on the endometrium. A. Diffuse placenta of the horse and pig. B. Cotyledonary placenta of ruminants. C. Zonary placenta of the dog and cat. D. Discoid placenta of the human and monkey.
A sterile heifer born twin to a normal bull calf is called a freemartin. This occurs when the female calf develops in the uterus with a normal male twin and they share the same blood supply (anastomosis of the placental blood vessels). When this occurs, the sex hormones from the earlier developing male twin pass across to the female twin, causing sexual differentiation of both male and female to proceed under control of male hormones. About 90% of heifer calves born twin to a bull calf are freemartins and can usually be detected clinically because of the shortened vagina (short advancement of a blunt instrument) and an enlarged clitoris.
Hormones
Pregnancy is maintained as a result of the predominance of progesterone. During gestation, progesterone is produced by the placenta and CL. The contribution from placental and luteal sources and the duration of their contribution varies among species. The CL source is needed by all species during early pregnancy, but is not needed by the mare and ewe after about 100 and 60 days, respectively. A CL is needed for most of pregnancy in the cow, bitch, and queen and for the entire pregnancy in the sow and doe. Although progesterone from the CL is not needed by the ewe, regression of the CL does not occur and luteal production continues, but placental production is dominant. Regression of the CL occurs in the mare about midway.
In the mare, endometrial cups begin to be formed at about day 35 of gestation within the endometrium from cells migrating from the placenta. The cups begin to secrete a hormone known as pregnant mare serum gonadotropin (PMSG), also referred to as equine chorionic gonadotropin (eCG), at about 35 days, which continues until about 130 days of gestation. Early pregnancy in the mare can be diagnosed by analyzing for PMSG. Pregnant mare serum gonadotropin helps to form new follicles, which ovulate and provide for additional corpora lutea. A greater supply of luteal progesterone is thereby ensured until the endometrial supply of progesterone is adequate for maintenance. In the mare, all corpora lutea regress by about 150 days. A newly identified hormone, dihydroprogesterone (DHP), now explains how horses sustain the last half of their pregnancies. It has been demonstrated in pregnant mares and through laboratory analyses that DHP is as potent as progesterone in activating the progesterone receptors of horses, triggering endometrial growth and maintaining pregnancy to term.
Diagnosis
It is often of economic importance to determine whether an animal is actually pregnant. Pregnancy is obvious during the late stages when the size of the fetus, uterus, and fetal fluids have increased to the point at which the abdomen has enlarged and definite dropping of the abdominal wall has occurred (known as bellying down). Rectal palpation is a useful procedure for detecting earlier signs of pregnancy, particularly in the cow. The hand is inserted into the rectum and structures located outside of the rectal wall can be felt.
Early pregnancy by rectal palpation in the cow is suggested if a corpus luteum is present and if one horn of the uterus is larger than the other. This condition can be apparent at 30 to 45 days. At about 3 months, the fetal membranes can be felt to slip away from the grasp when the uterus is lifted, and small caruncles in the uterine wall are palpable. Also at 3 months, a vibration or “buzzing” of blood in the uterine artery is palpable, known as fremitus. At 5 to 7 months the weight of the fetus causes the uterus to slip over the brim of the pelvis and the cervix becomes taut. The ovaries and fetus are difficult to palpate when this occurs because of their distance from the palpator, but definite caruncles are palpable.
After the fetus has descended over the brim of the pelvis in the cow, it may be possible to detect pregnancy by an external technique known as ballottement. Pressure is exerted on the lower right abdominal wall (see Figure 15-8) with the fist or knee in an inward and upward direction and then is released, causing the fetus to rise and fall in its suspending fluids. The fall should be felt by the manipulator.
The use of radiography for the diagnosis of pregnancy has had limited application in veterinary medicine. Penetration of the rays is restricted in large animals and exposure of the film is difficult. In small animals, such as the dog, exposure is adequate, but differentiation of a fetus is not possible until calcification of bones is adequate for contrast. This does not occur until about 45 days in the dog, and other means, such as palpation and observation, are often more useful for earlier diagnosis of pregnancy.
A biologic test for the detection of pregnancy can be performed in the mare based on the production of PMSG by the endometrial cups (see previous section). Historically, injection of serum taken from a mare at 40 to 130 days of pregnancy into a female rabbit that has been isolated from male rabbits for at least 30 days brings forth ovarian follicles that rupture and form reddened corpora hemorrhagica about 48 hours after injection. The corpora hemorrhagica can be seen when the rabbit is butchered or observed by other procedures when placed under anesthesia. Because the rabbit does not ovulate and form corpora hemorrhagica unless coitus occurs, only the injected PMSG could have caused the ovulation. Currently chemical and immunological tests (enzyme-linked immunosorbent assays (ELISA) for example) can be used to diagnosis pregnancy in the mare.
Ultrasonography is currently the most commonly used method for pregnancy diagnosis in small and large animals. The use of ultrasound for this purpose is being carried out by veterinary practitioners and theriogenologists. Transabdominal ultrasonography in the bitch is best used beyond 24 days of pregnancy where the amniotic vesicles are visible as black balls with a comma-shaped tissue mass within them. Beyond 24-30 days, beating hearts can be seen. In the queen, it is best used beyond 16 days of pregnancy and the amniotic vesicles are seen similar to their appearance in the bitch. Beyond 16-25 days, beating hearts can be seen.
Transrectal ultrasonography is used for large animals. An example for the cow indicates that the embryo is well demarcated by approximately day 18, with a highly accentuated curve in its anterior- posterior axis. Heartbeats are also visible at this time.
Human chorionic gonadotropin (HCG) is excreted in the urine of pregnant women. It is detectable about 8 days after ovulation, which is 1 day after implantation. Early detection of pregnancy in women is possible by diagnostic tests that use the presence of HCG in urine. Functionally, HCG is the signal for the corpus luteum to be maintained and thus sustain pregnancy.
■