Protein Function Depends on Protein Shape and Shape Changes
Protein function is founded on two molecular characteristics:
(1) proteins can bind to other molecules very specifically; and
(2) proteins change shape, which in turn alters their binding properties and their function.
The binding specificity of proteins is the result of their complex three-dimensional structure. Grooves or indentations on the surface of protein molecules, called binding sites, permit specific interactions with a molecule of a complementary shape, called the ligand. This complementary-shape mechanism underlying binding is similar to the shape interaction between a lock and key.Several aspects of the lock-and-key analogy are worth noting. As with a lock, only a small part of the protein is engaged in binding. The binding is very specific; and small changes in the shape of the binding site (keyhole) or the shape of the ligand (key) can cause major changes in protein (lock) behavior. Similar to the lock and key, the complementary- shape interaction serves a recognition function; only those molecules with the right shape affect protein function. This recognition function plays a primary role in information transfer. The protein recognizes a particular signal by binding to it, thus changing the protein’s shape and thus its function. Unlike the majority of locks, however, proteins frequently have multiple binding sites for multiple ligands.
Thus the three-dimensional shape of a protein, its conformation, determines protein function. A major force that stabilizes protein conformation is the hydrophobic interaction. Oily, “water-fearing” amino acids tend to congregate in the middle of a protein away from water, whereas hydrophilic ("water-loving”) amino acids tend to be found on the proteins outer surface interacting with the abundant cellular water. The hydrophobic interaction is also important in stabilizing the interaction of proteins with the lipids of biological membranes, as discussed shortly.
Protein shape is also stabilized by hydrogen bonding between polar amino acid pairs in the polypeptide (protein) chain.The same weak forces responsible for protein conformation are also used to hold the ligand in the protein-binding site. The position of the ligand in the binding site is stabilized by hydrogen bonds between the polar groups of the ligand and polar, amino acid side groups lining the binding site, just as hydrogen bonds within the polypeptide chain stabilize the shape of the polypeptide. Precisely because the same forces are responsible for the shape of the protein and for its binding properties, shape influences binding, and in turn, binding can influence protein shape. The ability of proteins to change shape is called allostery (Greek, "other shape”).
Allosteric changes in protein conformation arise in four general ways. One way, just mentioned, is that most proteins change shape depending on which ligands are bound at particular binding sites (Figure 1-1, A). The sequence— specific ligand binding → protein shape change → change in protein-binding properties and protein function —> this change regulates something—is a common molecular mechanism underlying physiological control. This method involves no alteration in the covalent structure of the protein.
A second method of producing conformational change, however, occurs as a result of the covalent modification of one or more of the amino acid side groups of the protein (Figure 1-1, B). By far the most common such change is the covalent addition of a phosphate group to the hydroxyl (—OH) group on the side chain of serine, threonine, or tyrosine residues in the protein. Because the phosphate group is highly charged, phosphorylation of a protein alters hydrogen bonding and other electrostatic interactions within the protein chain, altering its conformation and functional properties.
In a third method, some physiologically important proteins change shape in response to the electrical field surrounding the protein (Figure 1-1, C).
These respond to a voltage change by altering the position of charged amino acids, thus altering protein shape.The fourth method of protein shape change is the least well understood (not shown). Some proteins change shape in a controlled manner in response to mechanical forces. Although this is not surprising, because all solids and solidlike substances change shape at least slightly in response to force, we know relatively little about mechanosensitive proteins. The best current example is a protein involved in the very early events of hearing that changes its transport of ions in response to the mechanical stimulation of sound (small changes of air pressure in waves).
The significance of binding specificity and allostery can be better appreciated with two examples of their role in physiological function. The first example is the role of enzymes in synthesizing three small, structurally similar, nonprotein signaling molecules. This example shows how binding specificity is important in catalytic function and how allostery underlies the regulation of the synthesis. The second example is more complex: the role of proteins in the contraction of muscle. Ihe contraction of muscle shows how proteins can
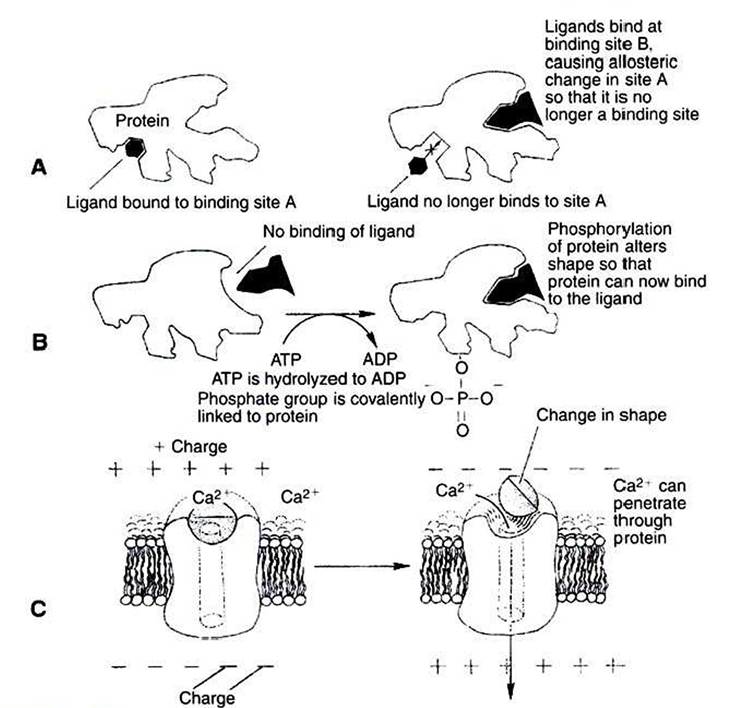
FIGURE 1-1 Three common mechanisms of allosteric shape change in proteins. A, Ligand binding. Ligand binding to an allosteric site (site B) on a protein changes the protein's conformation such that binding site A is altered; ligand no longer binds at site A because of the binding event at site B. Bz Phosphorylation. Addition of a phosphate group to a serine, threonine, or tyrosine residue of a protein alters the protein's conformation, changing its binding characteristics. In this hypothetical example, phosphorylation activates an otherwise inactive protein. Some proteins inactivate by this mechanism. AT^Adenosine triphosphate; ADPt adenosine diphosphate.
C, Voltage-dependent proteins.The conformation of some proteins, particularly ion channels, is altered by the electrical field surrounding the protein. Shown here is the opening (activation) of a voltage-dependent, gated Ca2* channel when the membrane depolarizes.
exploit the basic properties of specific binding and allosteric shape change to do more than one job of work at the same time; muscle proteins serve a structural role, serve a catalytic function, and couple the “downhill” hydrolysis of adenosine triphosphate (ATP) to do mechanical work, the “uphill” lifting of weight.