REGULATION OF VENTILATION
1. Where is the respiratory center for the regulation of ventilation located?
2. What is the response of the Hering-Breuer inspiratory-inhibitory reflex?
3. What is the response of the Hering-Breuer deflation reflex? What is its practical significance?
4.
What are the three humoral factors that influence ventilation?5. Where are the receptors located for the detection of O2 lack?
6. Why is H+ more influential as a stimulus to respiration than CO2 in the chemosensitive area of the brain stem?
7. Why is oxygen more important in the regulation of respiration in conditions such as pneumonia and pulmonary edema?
8. Why is there no increase in ventilation when there is O2 lack caused by carbon monoxide poisoning?
9. What is the braking effect? What is the example of how it can be observed?
Pulmonary ventilation is regulated closely to maintain the concentrations of H+, CO2, and O2 at relatively constant levels while meeting the needs of the body under varying conditions. If either the H+ or the CO2 concentration increases or if the O2 concentration decreases, their levels will be returned to normal by increasing ventilation. Conversely, if either the H+ or CO2 concentration decreases or if the O2 concentration increases, pulmonary ventilation will be decreased. This regulatory mechanism is controlled by changes in tidal volume, frequency of respiratory cycles, or both. The central mediator of these changes is the respiratory center in the brain stem, which has four specific regions (Figure 10-20):
1. Pneumotaxic center (PC): believed to modulate respiratory center sensitivity to inputs that activate termination of inspiration and facilitate expiration.
2. Apneustic center: believed to be associated with deep inspirations, such as the sigh.
3. Dorsal respiratory group (DRG): group of neurons predominately associated with inspiratory activity (particularly involved in lung inflation-induced termination of inspiration).
4. Ventral respiratory group (VRG): group of neurons containing inspiratory and expiratory neurons (assist in inspiration begun by those in the DRG and also provide for assisted expiration).
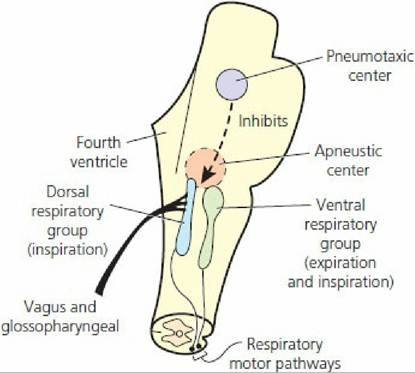
■ FIGURE 10-20 Components of the respiratory center. The pneumotaxic and apneustic centers are located in the pons and the dorsal and ventral respiratory groups are located in the medulla. (From Guyton AC, Hall JE. Textbook of Medical Physiology. 12th edn. Philadelphia, PA: Saunders Elsevier, 2011.)
A central pattern generator has been hypothesized; it is believed to be the neural network that provides for rhythmicity. This central pattern generator is also thought to be in the brain stem. It is influenced by inputs from the vagus and glossopharyngeal nerves and by chemoreceptors.
Neural Control
Impulses going to the respiratory center (afferent impulses) from several receptor sources have been identified. The Hering-Breuer reflexes are probably the most noteworthy. The receptors for these reflexes are located in the lungs, particularly in the bronchi and bronchioles. There are two components to the Hering-Breuer reflexes: (1) the inspiratory-inhibitory reflex or inflation reflex and (2) the inspiratory or deflation reflex. The nerve impulses generated by the receptors of the Hering-Breuer reflexes are transmitted by fibers in the vagus nerves to the respiratory center. The effect of inflation-receptor stimulation (stimulation of neurons in the DRG) is to inhibit further inspiration and to stimulate expiratory neurons in the VRG. The inspiratory or deflation reflex component is activated at some particular point of deflation. The deflation receptors might not be activated to bring about the next inspiration during eupnea, but they might be active when deflation is more complete.
The deflation-receptor reflex stimulation can be elicited in anesthetized dogs by manual compression of the thorax, which is followed immediately by inspiration. Practical use of this reflex is appropriate for respiratory depressed or unresponsive animals to promote more adequate ventilation in the former or to initiate ventilation in the latter. During exercise when tidal volume and frequency are increased, it would appear that the deflation reflex becomes more active in order to hasten the beginning of the next inspiration.In addition to lung receptors, there are other peripherally located receptors that modify the basic rhythm. Stimulation of receptors in the skin is excitatory to the respiratory center, and deeper than usual inspiration can be noted. Their excitation to the inspiratory area might be through the apneustic center, inasmuch as inspiratory gasps are occasionally noted. Advantage is taken of these receptors when breathing stimulation is desired in newborn animals. Rubbing the skin with a rough cloth often initiates the breathing cycles. An assist to ventilation needed during muscle activity is obtained from receptors located in tendons and joints. They will be stimulated when muscle contraction causes movement. It is also believed that when impulses are directed to skeletal muscles from the cerebral cortex, collateral impulses go to the brainstem and stimulate the respiratory center to increase alveolar ventilation. This mechanism might account for increases in ventilation that are not explainable by mere observation of changes in carbon dioxide, oxygen, and hydrogen ion concentration in the blood.
Several respiratory reflexes originate from receptors in the upper air passages. Stimulation of the mucous membranes in these regions causes reflex inhibition of breathing. A striking example of this reflex is the inhibition of breathing that occurs in diving birds and mammals whereby reflex inhibition of breathing is observed when they submerge. Similarly, sneezing can be observed after stimulation of the nasal mucous membrane by various mechanisms.
The function of these latter reflexes is protection of the delicate respiratory passages and the alveoli of the lungs from harmful substances (e.g., irritating gases, dust, smoke, food particles) that might otherwise be inspired. Stimulation of the laryngeal mucous membrane in the unanesthetized animal causes not only inhibition of breathing but also usually powerful expiratory efforts (coughing). To ensure protection, the glottis is closed and the bronchi may be constricted. Endotracheal intubation is often difficult in lightly anesthetized animals because of reflex closure of the glottis.Ordinary respirations proceed involuntarily. It is generally true, however, that they can be altered voluntarily within wide limits - they can be hastened, slowed, or stopped altogether, for a while. Phonation and use of the abdominal press in the expulsive acts.of defecation, urination, and parturition are all examples of (more or less) complete voluntary control of the respiratory movements. These acts, however, are not concerned with gas exchange between the organism and its environment, but represent secondary functions of the respiratory apparatus.
Afferent impulses from pressure receptors in the carotid and aortic sinuses have as their principal function a role in the regulation of circulation, but impulses from these receptors also go to the respiratory center. The impulses are inhibitory in nature - the higher the blood pressure, the greater the inhibition to respiration. Because of the influence of inspiration on return of blood to the heart, it can be seen that the reduction in inspirations would slow down the return flow of blood to the heart and thus help to lower blood pressure.
Humoral Control
Humoral control refers to those factors in the body fluids that influence ventilation: carbon dioxide, hydrogen ions, and oxygen. Because these are constituents of the body fluids, it seems natural that they should exert the greatest influence on ventilation in maintaining constancy.
Their concentrations in the blood affect alveolar ventilation in several ways:1. Carbon dioxide increase causes alveolar ventilation to increase; its decrease causes alveolar ventilation to decrease.
2. Hydrogen ion increase causes alveolar ventilation to increase; its decrease causes alveolar ventilation to decrease.
3. Oxygen increase causes alveolar ventilation to decrease; its decrease causes alveolar ventilation to increase.
Influences of Carbon Dioxide and Hydrogen Ions
The effects of carbon dioxide and hydrogen ions are mediated through bilateral chemosensitive areas beneath the ventral surface of the medulla (Figure 10-21). Because of the much greater diffusibility of carbon dioxide, as compared with H+, it is distributed more quickly from the blood to the interstitial fluid of the medulla and to the cerebrospinal fluid than hydrogen ions. It is believed, however, that the H+ concentration of the interstitial fluid of the brain stem is the deciding stimulus for respiratory drive. The influence of CO2 is exerted by its conversion to H+ through the hydration reaction (Equation 10-1; see previous section).
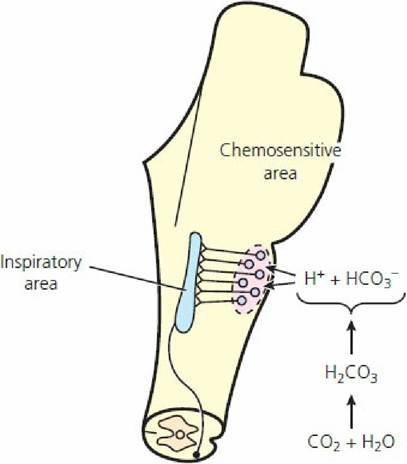
■ FIGURE 10-21 The chemosensitive area of the brain stem respiratory center. The chemosensitive area is stimulated by hydrogen ions, which are formed by the conversion of carbon dioxide through the hydration reaction. (From Guyton AC, Hall JE. Textbook of Medical Physiology. 10th edn. Philadelphia, PA: WB Saunders, 2000.)
Influences of Oxygen
The influence of oxygen is transmitted from the carotid and aortic bodies to the respiratory center. The carotid and aortic body receptors also respond to carbon dioxide and hydrogen ion concentration, but the effectiveness of the carotid and aortic body response to carbon dioxide and hydrogen ions is far less than the response from the brain stem. Thus, the carotid and aortic bodies are considered to be the most influential for the regulation of oxygen.
These bodies are distinct structures with an abundant blood supply located just outside the aortic arch, at the division of the carotid arteries. They respond to changes in the PaO2 of blood. Blood with reduced amounts of hemoglobin, and consequently less oxygen, has the same PaO2 as blood with normal hemoglobin and oxygen (see Figure 10-17), and thus no ventilation response would be elicited because there is no change in PaO2. Also, blood in which oxygen has been displaced from hemoglobin by carbon monoxide has the same PaO2 as normal blood (oxygen in solution) and there would be no increase in ventilation. The PaO2 would remain the same because it is an expression of alveolar PO2 (which has not changed) and represents the PO2 of oxygen in solution. In the case of decreased hemoglobin (e.g., as in anemia) ventilation might be increased, not because of less oxygen, but because of greater hydrogen ion concentration caused by reduced buffering associated with the hemoglobin decrease. In the case of carbon monoxide poisoning and lack of oxygen carried by hemoglobin, ventilation is not increased, not only because the PaO2 is normal but also because there is adequate hemoglobin present for buffering hydrogen ions.Arterial blood PO2 must be in the range of 30 to 60 mm Hg for the respiratory center to receive stimulation to ventilation from the carotid and aortic bodies (Figure 10-22). This seems to be an appropriate range, because hemoglobin is still about 90% saturated with oxygen at a PO2 of.60 mm Hg. Also, the slowing effect of an increased arterial PO2 is subtle and would not normally be observed in animals breathing atmospheric air because the arterial PO2 seldom increases above 100 mm Hg. The slowing effect is noted, however, in anesthetized animals breathing an oxygen-enriched atmosphere, in which the arterial PO2 could increase to 350 to 400 mm Hg (Figure 10-23).
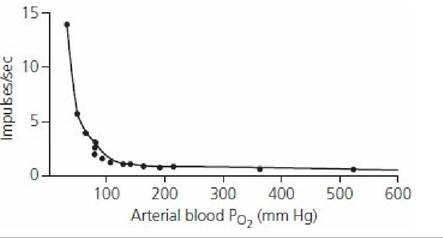
■ FIGURE 10-22 Effect of arterial oxygen partial pressure on the number of impulses per second from the carotid body to the respiratory center. The impulses are excitatory. (From Reece WO. Respiration in mammals. In: Reece WO, ed. Dukes’ Physiology of Domestic Animals. 13th edn. Ames, IA: Wiley-Blackwell, 2015.)
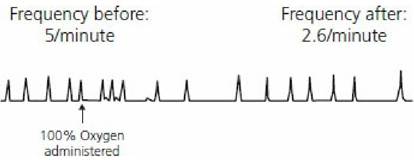
■ FIGURE 10-23 Pneumogram showing the effect of oxygen enrichment on respiratory frequency. An oxygen atmosphere was provided to a pentobarbital-anesthetized dog. Note the decreased respiratory frequency after administration of oxygen (drawn from actual recording). (From Reece WO. Respiration in mammals. In: Reece WO, ed. Dukes’ Physiology of Domestic Animals. 13th edn. Ames, IA: Wiley-Blackwell, 2015.)
Importance of Oxygen Regulation
The regulation of ventilation by oxygen is not ordinarily thought to be important. There is usually no problem in maintaining arterial blood PO2 in the range of 80 to 100 mm Hg, and it is not advantageous to have it higher than 100 mm Hg because hemoglobin is almost saturated at that partial pressure. Ventilation could even be reduced to about 50% of normal and hemoglobin would still be considerably saturated. Accordingly, the most important chemical factor in the regulation of ventilation is the concentration of carbon dioxide; relatively small changes can have an effect. The regulation of ventilation by oxygen becomes more important in such conditions as pneumonia and pulmonary edema, in which gases are not diffused as readily through the respiratory membrane.
Decreased diffusion is more noticeable for oxygen than for carbon dioxide (see previous section) because of the smaller diffusion coefficient for oxygen. Hyperventilation caused by oxygen lack can therefore reduce the carbon dioxide concentration (because CO2 readily diffuses) and thus the consequent reduced formation of hydrogen ions (see Equation 10-1) so that they become ineffective in stimulating increased ventilation. The oxygen deficiency mechanism (originating from the carotid and aortic bodies) continues to function and provides the drive to increase ventilation.
Braking Effect
The effect of decreased concentrations of H+ and CO2 and increased concentration of O2 to slow down ventilation is referred to as a braking effect. The braking effect of O2 was shown to be unimportant, but it is important for CO2 and H+ to decrease ventilation because they are both involved in maintaining the acid-base equilibrium of the body fluids. The uncontrolled lowering of either CO2 or H+ would result in some degree of alkalemia (decreased H+ concentration in blood). A braking effect can be observed when anesthetized animals being hyperventilated with a respirator are removed suddenly from the respirator. A minute or more might be required for CO2 and H+ to accumulate to a level at which they no longer exert their braking effect and breathing finally resumes. In this example, oxygen lack is apparent, and it could also be a contributory factor in the resumption of breathing.
The factors that influence ventilation are summarized in Figure 10-24.
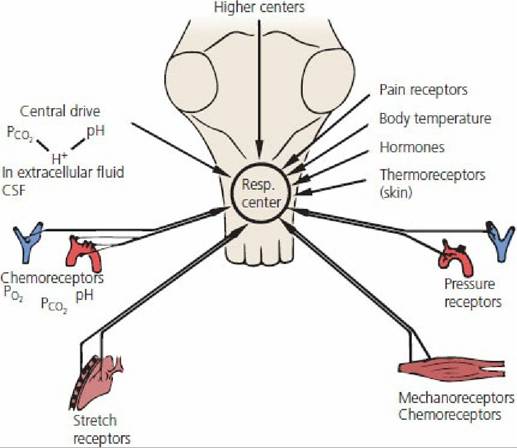
■ FIGURE 10-24 Summary of factors that influence pulmonary ventilation. (From Schmidt PS,
Thews G, eds. Human Physiology. Berlin: Springer-Verlag, 1989.)