RESPIRATORY SYSTEM
Apart from gas exchange, the lizard uses its lungs for threat and display, buoyancy, escaping from predators and vocalization. It achieves this by having large volume, highly compliant lungs that can easily be inflated by gular pumping (Perry & Duncker 1978; Wood & Lenfant 1976).
Upper respiratory tract
The lizard palate has two long slit-like openings rostrally where the internal nares and Jacobson’s organs open into the mouth and this can be a common site for discharges to develop. The glottis is variable in location and can be found very rostrally or at the back of the tongue, depending on the species. It is normally closed except during inspiration and expiration (Fig. 4.16).
Lower respiratory tract
The trachea has incomplete tracheal rings and bifurcates in the thoracic cavity near the heart. As they have no diaphragm, lizards breathe by expansion and contraction of the ribs. All three structural lung types are found in lizards: unicameral, paucicameral and multicameral (Fig. 4.17).
Unicameral lungs
Primitive lizards (e.g., the Green lizard, Lacerta viridis) lack any division between the pleural and peritoneal cavities. The heart lies in the pectoral inlet and the lungs form extensive hollow sacs that consist of a single chamber, hence “unicameral” (Fig. 4.18). They generally occupy the cranial part of the pleuroperitoneum but in some lizards like skinks (Scincidae) they have caudal non-respiratory dilatation, similar to airsacs. These airsacs are poorly vascularized and so can be a site of infection (Perry 1989; Perry & Duncker 1978).
Paucicameral lungs
Intermediate species have developed a membrane like a postpulmonary septum that connects to the pericardium. These paucicameral lungs have finer partitioning, with some having large caudal dilatations like airsacs (Fig. 4.19). In species like the chameleons these can be inflated to increase lung volume by over 40% in order to ward off predators (Perry 1989; Perry & Duncker 1978).
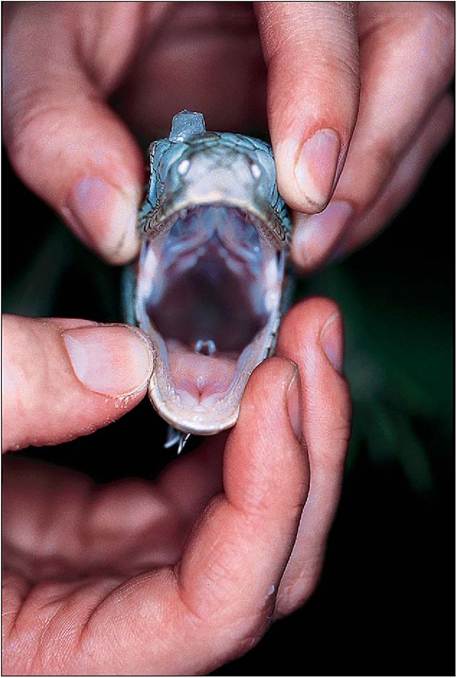
Figure 4.16 • Open mouth view of lizard showing open glottis. Respiration in reptiles is triphasic with the pause phase being the longest.
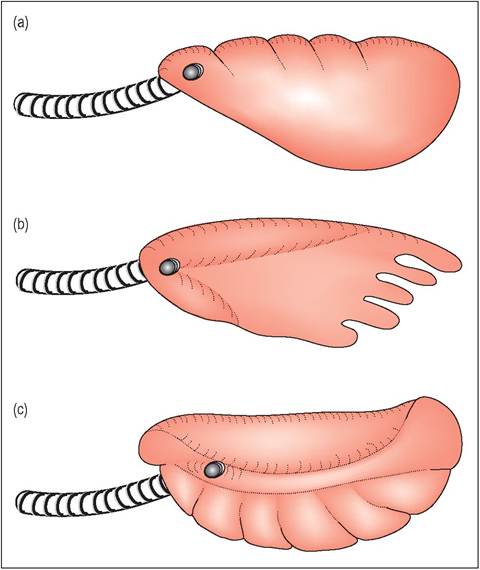
Figure 4.17 •
(a) Unicameral lung of simple lizard (e.g., Green lizard, Lacerta viridis)
(b) Paucicameral lungs with a few simple divisions. In some lizards, like the Common chameleon (Chamaeleo chameleon) there are airsac dilatations caudally.
(c) Multicameral lungs are more advanced multichambered lungs with a single intrapulmonary bronchus.
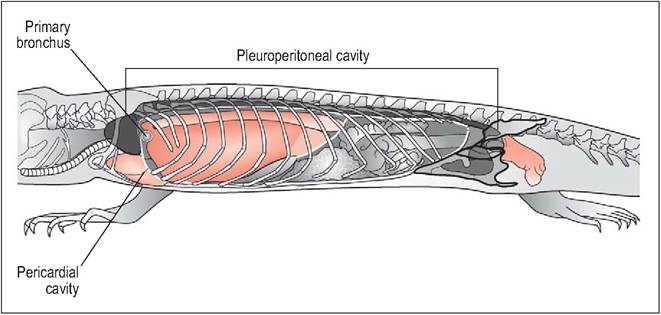
Figure 4.18 • Unicameral lizard - schematic diagram of celomic cavities (left lung, liver lobe and stomach removed). In primitive lizards, the lack of a postpulmonary septum results in their having an extensive combined pleuroperitoneal cavity. The heart is located cranially in the pectoral region.
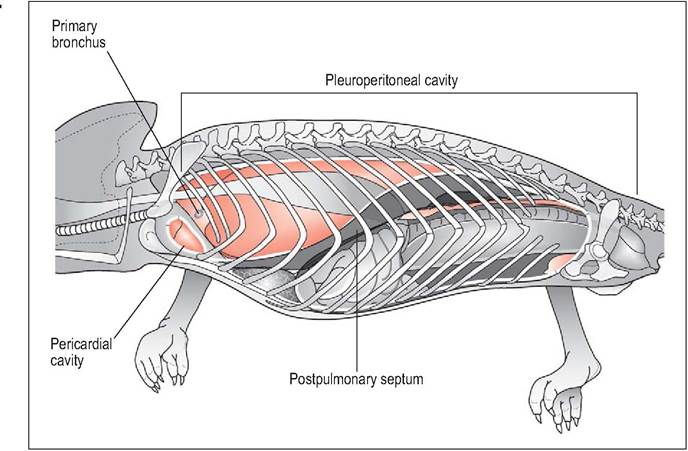
Figure 4.19 • Paucicameral lizard - schematic drawing (left lung, liver lobe and stomach removed) of celomic cavities showing small postpulmonary septum and partial separation between pleural and peritoneal cavities. The heart is located in the pectoral region.
Multicameral lungs
In the more advanced monitor lizards the postpulmonary septum completely divides the lungs in the pleural cavity from the peritoneal cavity. The lungs more closely resemble the lungs of primitive mammals and occupy the cranial dorsal and ventral cavity. As they have expanded cranioventrally the heart now occupies a more caudal position on the mid sternum rather than at the pectoral inlet. Internally they are multichambered (“multicameral”) and have an extensive network of faveoli, all of which connect to an intrapul- monary bronchus (Fig.
4.20). Large volume and highly compliant lungs give an increased tidal volume and low respiratory rate (Bellairs 1969d; Bennett, AF 1972; Perry 1989; Perry & Duncker 1978).Respiration cycle
Both inspiration and expiration are active processes that are followed by a non-ventilatory portion of varying length (up to 30 minutes in some species) (Wood & Lenfant 1976). Respiration is based on the negative celomic pressure that is produced by the intercostal muscles, aided by the trunk and abdominal muscles. In addition the lung walls contain smooth muscle that contracts and relaxes.
Like all reptiles, lizards have little capacity for aerobic respiration and switch to anaerobic quite fast; hence, they cannot sustain activity for long but work better in bursts. However, the large Australian Sand monitor (Varanus gouldii), has twice the aerobic capacity of other reptiles due to its highly developed lung falveoli and more myoglobin. This means it can sustain vigorous activity for longer without getting fatigued (Bennett, AF 1972).
CLINICAL NOTE
Lizards use the same intercostal muscles for breathing as for locomotion and so they actually cease breathing while running and then stop to breathe, which may explain why they are often active in short bursts (Pough I998c).
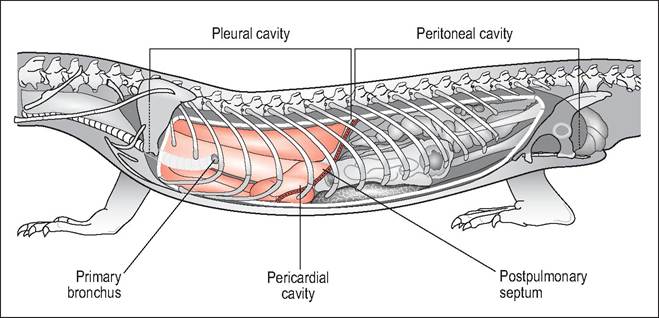
Figure 4.20 • Multicameral lizard (Monitor lizards) - schematic drawing showing complete separation of pleural from peritoneal cavities. This has allowed the lungs to expand cranially and ventrally causing the pericardial sac and heart to move caudally into the mid celom.
DIGESTIVE SYSTEM
The majority of lizards are omnivorous and carnivorous; only about 40 species are herbivorous and these tend to be larger in body size.
Prehension of food
Carnivorous lizards ingest their prey in two ways: small prey are seized and crushed by the jaws. Large prey are ingested by what is called inertia feeding.
This is where the prey is lifted off the ground and shaken violently and then the jaws relax and slide forward over the inert prey item (Pough 1998e).Dentition
Unlike snakes and chelonians, lizards do chew their food and will tear off pieces if the food item is too big to swallow. Lizard teeth are simple conical structures that have no sockets and are regularly shed and replaced in waves to always ensure there are adequate teeth for prehension of food. The replacement tooth is formed in the mucosa of the base, rather than below it as in mammals (Edmund 1970; King 1996b). Lizards can have either pleurodont or acrodont teeth (Fig. 4.21).
Pleurodont
Pleurodont teeth are attached to the inner sides of the mandible and are found in iguanids and varanids. These teeth are continually shed and replaced by new teeth forming at the lingual side of the base. The gingival margin is just behind the bony crest supporting the teeth (Edmund 1970; King 1996b).
CLINICAL NOTE
Iguanas have a rapid rate of teeth replacement. Each quadrant of their mouth has about 20 to 30 teeth and these are replaced up to five times a year, so a medium-sized iguana can replace 500 teeth a year!
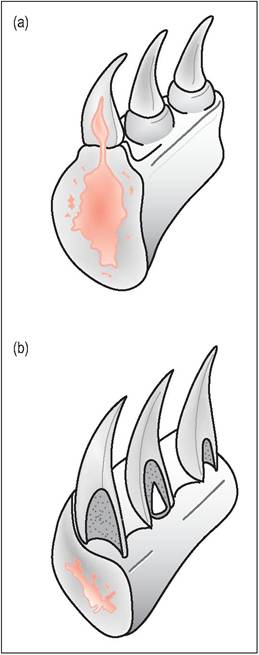
Figure 4.21 • Lizard dentition.
(a) Acrodonts, such as the agamids and chameleons, have the teeth attached to the biting edges of the mandible and maxilla.
(b) Pleurodonts, such as the iguanids and varanids, have the teeth attached to the inner sides of the mandible.
Acrodont
Acrodont teeth are attached to the biting edges of the mandible and maxillae and are found in agamids and chameleons. They are not replaced throughout life and wear with age, leaving the jaw to act as a cutting edge in the end. In most acrodont species (with the exception of the chameleon) the most rostral teeth are pleurodont (McCracken 1999).
Egg tooth
Newly hatched lizards have an egg tooth on the rostrome- dial side of the premaxilla.
This is a modified premaxillaryCLINICAL NOTE
The gingival margin lies further back from the teeth in acrodont lizards, making species like Bearded dragons (Pogona vitticeps) and Water dragons (Physignathus spp.) more prone to periodontal disease if fed an inappropriate diet (McCracken 1999).
tooth and helps to slit the eggshell during hatching. Many species of gecko have a double egg tooth (Edmund 1970).
Venom
There are only two species of poisonous lizard: the Gila monster (Heloderma suspectum) and the Mexican Beaded lizard (Heloderma horridum), which come from the southwestern US and Mexico. Unlike snakes, the venom is located in large sublingual glands in the lateral lower jaw. The venom is secreted via ducts into the labial side of the mandible where adjacent grooved teeth transfer the neurotoxic venom (Barten 1996; Bellairs 1969c; Pough 1998e).
Tongue
The tongue is mobile and protrusible and attached to the hyoid apparatus at its base. It functions to bring scent particles to Jacobson's organ for olfaction, and for lapping, swallowing, and moving food through the oral cavity. Taste buds are poorly developed. In chameleons it is used for food prehension and the sticky tip can be projected rapidly over half the length of its body to catch prey (Liem et al. 2001). In monitors and tegus it is deeply forked and is used for exploring the environment, as in snakes. The Leopard gecko uses its mobile pink tongue to clean its corneas after feeding (Evans 1986).
CLINICAL NOTE
In the Green iguana the rostral tip of the tongue is a darker pink color but this should not be mistaken for a pathological lesion (Barten 1996).
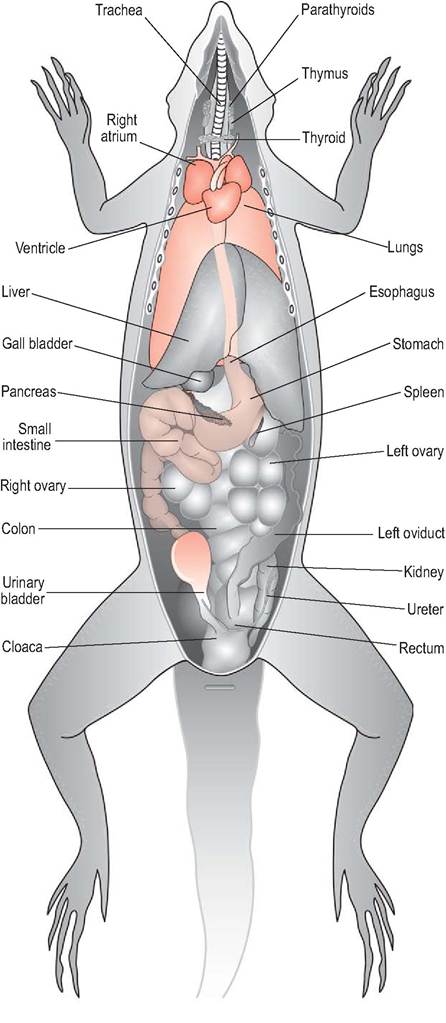
Figure 4.22 • Ventral view of female lizard showing internal anatomy.
Gastrointestinal tract
The gastrointestinal tract of insectivores and omnivores is relatively simple (Figs. 4.22 and 4.23). A short, thin-walled esophagus enters the stomach on the left side of the abdomen.
The stomach is tubular and simple and leads into a short small intestine and large intestine. Unlike the avian gizzard the swallowing of stones to aid digestion is not normal behavior in lizards. Some species like geckos use their tail for fat storage as an emergency energy source (Fig. 4.24).CLINICAL NOTE
Lizards drink with their tongue, putting it slowly in and out of water, whereas snakes prefer to immerse their head in water and gulp it down.
Herbivores
Only about 3% of lizards are herbivorous (King 1996b) and most are found in the families Iguanidae (e.g., Green iguana)
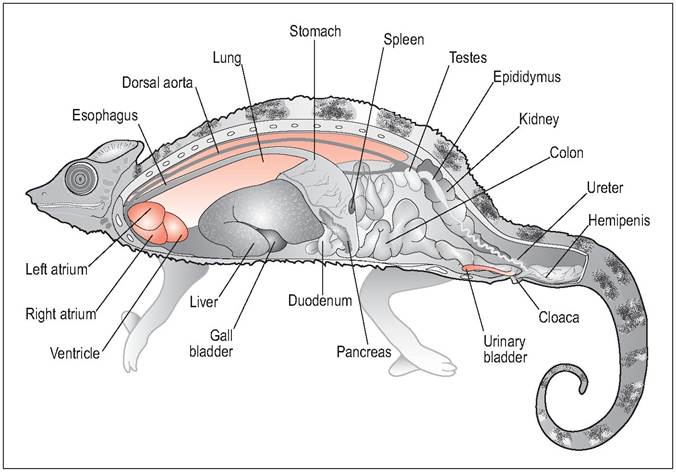
Figure 4.23 • Lateral, male chameleon.
midsagittal view of
and Agamidae (e.g., Uromastyx). Many herbivores have blunt heads like tortoises, giving them powerful jaw musculature for mastication. Both pleurodont and acrodont teeth can occur. Unlike insectivores, which eat bite-sized insects, herbivores have to crop down pieces of vegetation and so use their teeth as if they were scissors. However, unlike in herbivorous mammals, chewing on one side cannot take place as the upper and lower jaws are the same width (King 1996b).
Herbivores such as the iguana are hindgut fermenters and are really only found in tropical areas where high ambient temperatures facilitate fermentation. The small intestine is short and the large intestine (cecum and colon) takes up 50% of the length. Protein and digestible nutrients are absorbed in the stomach and small intestine while the indigestible cellulose is broken down and absorbed as volatile fatty acids in the cecum and colon. Most herbivores have a large partitioned proximal colon which increases surface area and slows the passage of ingesta, thus giving more time for hindgut fermentation. The iguana has large transverse folds in the proximal colon (Fig. 4.25), subdividing it into five pockets which decrease in size distally, and can retain food there for up to 3½ days (King 1996b, 1996c; Troyer 1984).
| Table 4.3 Examples of herbivorous lizards | |
| Common name | Scientific name |
| Green iguana Marine iguana Common chuckwalla Solomon Island skink Spiny tailed lizard (Dab) | Iguana iguana Amblyrhynchus cristatus Sauromalus ater Corucia zebrata Uromastyx acanthinurus |
Newly hatched iguanas have no microbial flora and in the wild they populate their gut by eating adult feces. They selectively eat more digestible proteins and have shorter transit times in order to gain enough energy for rapid growth (Troyer 1984).
Herbivorous species rely on high ambient temperatures and microorganisms to aid fermentation of cellulose into VFAs (King 1996b, 1996c). Total transit time is very slow in comparison to other non-ruminant herbivores. Mammals, from rabbits to elephants, take around 68 hours in comparison to up to 140 hours in the lizard. This is because reptiles do not ferment food during night cooling and have a much slower metabolism (Troyer 1984). It is also inversely related to body temperature (King 1996c).