A Second Line of Defense Consists of Phagocytic Cells of the Myeloid and Macrophage-Monocyte Lineages
Should an antigen survive the first line of the body’s defense (i.e., body surfaces) and penetrate into blood vessels and tissues, the body’s defense relies on cellular response.
Key cells involved in cellular defense are phagocytic cells, which are an integral part of innate immunity. These cells, based on their cellular origin, are broadly divided into myeloid and macrophage-monocyte lineages. Included in the myeloid lineage are neutrophils, eosinophils, and basophils. The macrophage-monocyte series includes monocytes and macrophages. Neutrophils constitute the largest percentage of white blood cells in most species (60%-65%), except in ruminants (20%-25%). Neutrophils have a short life span in the blood (half-life, ~12 hours), but in tissues their longevity increases to several days. Neutrophils are approximately 12 μm in diameter, with Inultilobulated nuclei and a cytoplasm rich in granules. The population of granules is composed of both primary and secondary granules. The primary granules contain important bactericidal enzymes such as myeloperoxidase, lysozymes, acid hydrolases (e.g., β-gkιcuroπidase, cathepsin),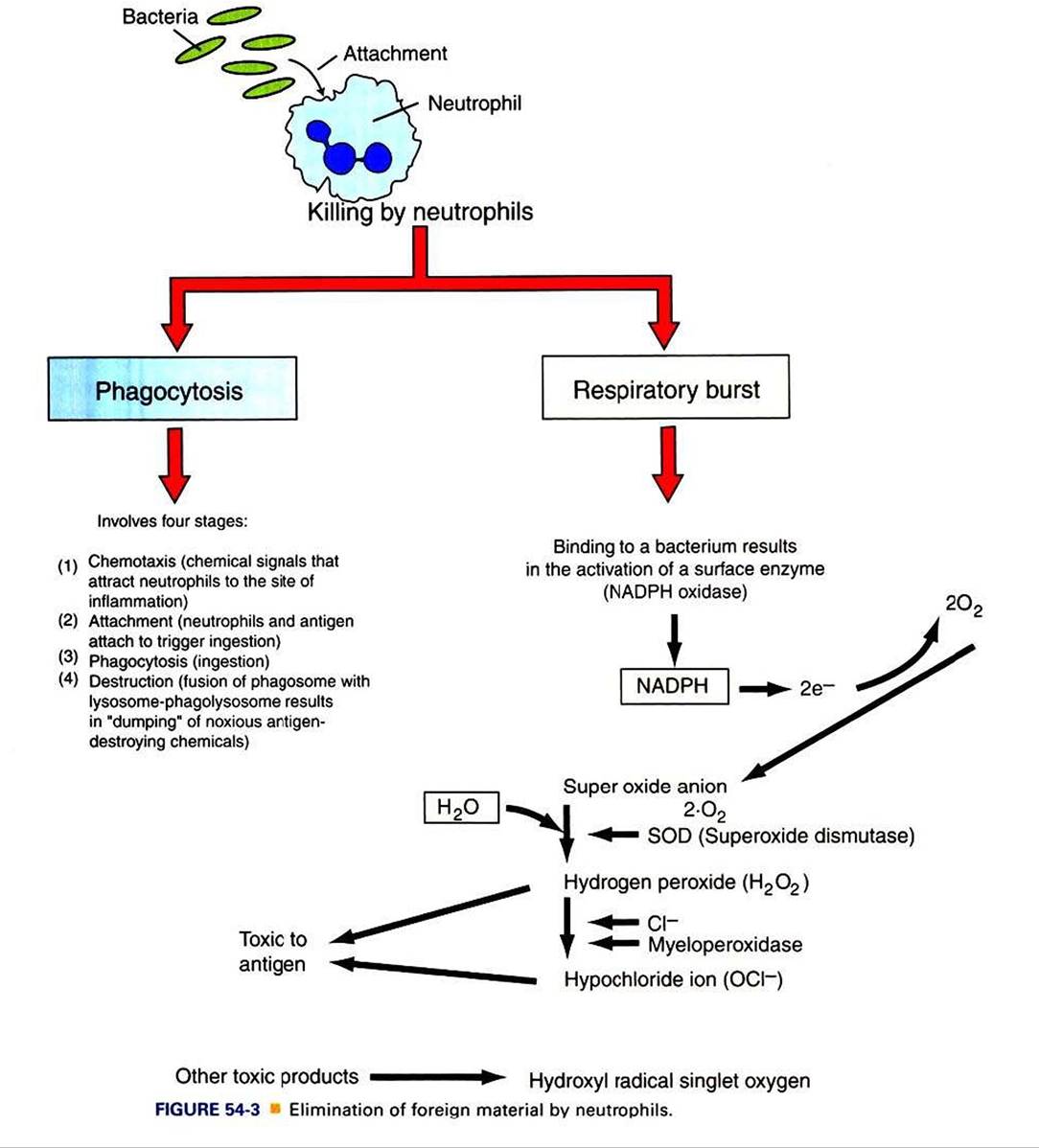
and neutral proteases against elastase hydrolases. Neutrophils also have defensinst small proteins that are inserted between the lipid bilayers and disrupt the interactions emanating from lipid membranes. The secondary granules include lysozymes, lactoferrin, and collagenases.
Neutrophils are considered the first responder cells to combat invading antigens. The primary function of neutrophils is to capture and destroy antigens. Neutrophils, unlike monocytes or macrophages, respond rapidly to invading antigens and readily phagocytose antigens. However, neutrophils lack the capacity to present antigens to lymphocytes.
Neutrophils destroy antigens by two different but complementary mechanisms: (I) phagocytosis and (2) respiratory burst. Phagocytosis in turn is divided into four arbitrary stages: (a) chemotaxis, (b) adherence or attachment, (c) phagocytosis, and (d) destruction (Figure 54-3).Neutrophils are attracted to the site of infection in tissues by chemokines and chemical messengers that are released when tissues are damaged. In response to chemical signals and chemokines, vascular endothelial cells induce the expression of adhesion molecules. Neutrophils bind to these cellular adhesion molecules (CAMs) through specific receptors and are triggered to leave the circulation by crossing capillary walls (diapedesis) into tissues. Once neutrophils move out of the circulation, they move toward the antigen. The contact between neutrophils and antigen is greatly facilitated when antigens are coated or bound by a host’s proteins, such as complement or antibodies. These proteins that enhance contact and phagocytosis by neutrophils or other phagocytes are called opsonins. The contact of neutrophils with antigens triggers infolding of the cell membrane (by the action of actin and myosin), and the antigen is trapped in a vacuole called a phagosome. The primary granules move toward the phagosome and fuse their membrane to become phagolysosomes, and in the process the granules release deleterious bacteriostatic and bactericidal biomolecules. Thus, in the contained environment of phagolysosomes, the antigen is destroyed.
A concurrent mechanism by which neutrophils kill invading microbial antigens involves respiratory burst (see Figure 54-3). On contact of neutrophils with an antigen, consumption of oxygen is immediately increased, 70- to 100-fold. This results in activation of an enzyme, NADPH oxidase, which forms an electron transport chain with cytosolic NADPH as an electron donor of oxygen. A molecule of oxygen accepts two donated electrons to result in a superoxide anion (O2^).
This O2", under the influence of the enzyme superoxide dismutase and in the presence of water, will chemically react to yield hydrogen peroxide (H2O2), which is toxic to microbes. This H2O2, under the influence of myeloperoxidase and utilizing chloride ions (CΓ), catalyzes oxidative reactions to form H2O2 and halide ions. All these products are highly toxic to the antigens.Neutrophils are also known to release hydroxyl radical singlet oxygen, which is toxic to bacteria.
Neutrophils have limited energy and a relatively short life. Elastases and collagenases released from these dying neutrophils serve as a powerful chemoattractant for another group of phagocytes called macrophages, and therefore neutrophils are sometimes referred to as “martyrs of the immune system.” Macrophages are attracted by bacterial products as well but also through the chemotactic factors released from damaged tissues. Macrophages differ from neutrophils in several important aspects. Macrophages, even though not rapid-responder cells, have extensive ability to phagocytose antigens repeatedly. These long-lived cells secrete large quantities of cytokines and chemokines that play a key role in regulating immune responses. Some of these cells even have the ability to present antigens to the immune system. Macrophages are present throughout body tissues where entry of antigens is likely.
Macrophages are round or elongated cells and express many surface receptors that include major histocompatibility complex antigens I and II (MHC class I and II). MHC classes I and II play a major role in antigen recognition and presentation (see chapter 55). Macrophages differ in their morphology based on tissues and thus are called by different names. For example, in the lymphoid organs, these cells are macrophages, whereas in the liver they are known as KupffePs cells (Figure 54-4). Macrophages are derived from the bone marrow hematopoietic cells and are initially called monoblasts, which mature and move into circulation and are known as monocytes.
When monocytes move into the tissues, they are called macrophages. Macrophages are larger than neutrophils and are rich in rough endoplasmic reticulum and Golgi bodies, indicating their extensive ability to produce and secrete immUnoregulatory proteins.Ingestion of antigen will activate the metabolic machinery of macrophages, including increasing lysosomal and bactericidal activity and Upregulating the inducible nitric oxide
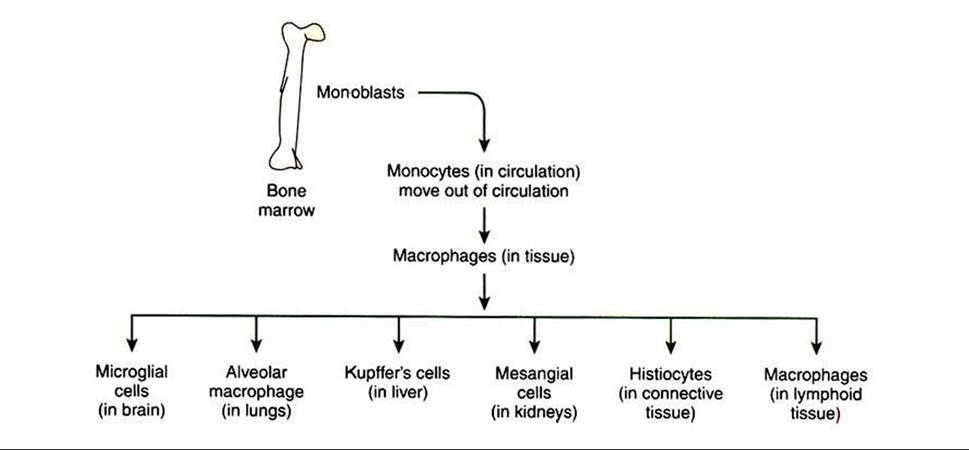
FIGURE 54-4 Anatomical location of macrophages throughout the body.
synthase gene (iNOS) that encodes for iNOS protein, which in turn is responsible for enhanced release of potent antimicrobial nitric oxide. Macrophages also release several oxygen free radicals that are also antimicrobial. Thus, macrophages aid in killing of antigens by phagocytosis and by respiratory burst (oxygen and nitrogen free radicals) (see Figure 54-3). Macrophages can potentially secrete more than 100 proteins. Some of these proteins, such as interleukin 1 (IL-1), interleukin-12 (IL-12), tumor necrosis factor alpha (TNF-α), interleukin 18 (IL-18), and interleukin 27 (IL-27), play a central role in activating lymphocytes, especially naive T lymphocytes (Figure 54-5). After an effective immune response is initiated, macrophages also secrete cytokines such as interleukin 10 (IL-IO) and transforming growth factor beta (TGF-β) which downregulate the immune response.
Once the antigen is cleared by macrophages, these cells also play an important role in repairing the damaged tissues. Macrophages secrete angiogenic factors to enhance the blood supply. For example, IL-I secreted by macrophages stimulates fibroblasts to secrete collagen to rebuild tissues.