Secondary and Tertiary Active Transport Mechanisms Utilize the Transcellular Sodium Ion Electrochemical Gradient as Their Source of Energy
)ust as a large stone resting atop a hill represents potential energy, so does the electrochemical gradient of sodium ions (Na ) across the enterocyte membrane. Gravity imparts potential energy to the stone, whereas diffusion forces impart potential energy to Na* outside cells.
Transport mechanisms that harness the potential energy of the sodium gradient are referred to as secondary active transport. Various transport pathway proteins exist for secondary active transport.One type is referred to as a co-transport protein or symport. The characteristic of a co-transport protein is that it has binding sites for one or more Na' ions as well as an additional binding site for some other specific molecule. For example, the glucose co-transport protein has one binding site for glucose and two for Na'. Co-transport proteins exist in the apical membrane of enterocytes. When the binding sites are unoccupied, they face the intestinal lumen. When all binding sites are occupied, a change in molecular configuration results
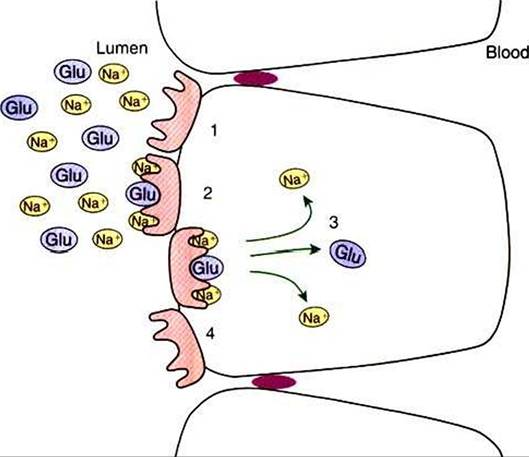
FIGURE 30-12 Co-transport is made possible by the allosteric transformation of transport proteins that lie in the apical membrane.The co-transport protein has two binding sites for sodium ions (NaV and one for glucose (GIu). When all three binding sites are occupied, the protein changes configuration in such a way as to transport the three ligands into the cell.The favorable gradient for sodium movement is maintained by the continuous action of the N a’,K4-ATPase pump (see Figure 30-13).
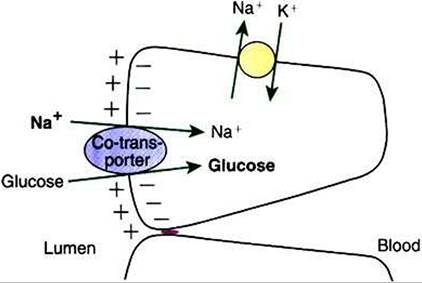
FIGURE 30-13 During co-transport, glucose is transported against an unfavorable concentration gradient.This diagram illustrates that the large sodium concentration difference across the apical membrane provides energy to transport glucose against its concentration gradient.The sodium concentration gradient, created by the action of the Na+,K* ATPase pump, provides energy to drive this reaction.
in the translocation of binding sites, with their ligand molecules, to the interior of the cell. When this happens, the Na+ ions along with the co-transported molecule are released into the intracellular fluid. Thus, there is transport of sodium and another molecule, such as glucose, across the apical membrane. When the ligand molecules are released, the protein assumes its original configuration so that the binding sites are again on the extracellular surface of the apical membrane, ready to transport additional molecules (Figure 30-12).
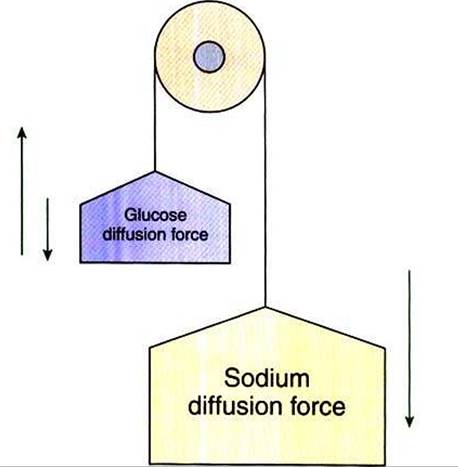
FIGURE 30-14 * Secondary transport is an important concept. The tremendous sodium concentration difference between intracellular and extracellular fluid could be likened to the force of gravity; a pervasive force that affects many relationships in our environment. Movement of most ions, glucose, and many other organic molecules across the intestinal epithelium is driven by the strength of the sodium concentration difference.
This process proceeds only as long as there is an electrochemical gradient for the Na*. When this gradient is large, as is normally the case, it can provide the energy to “pull” the co-transported molecule, such as glucose, from an area of lower concentration to one of higher concentration, as illustrated in Figures 30-13 and 30-14. Although the movement of a molecule against its concentration gradient represents expenditure of energy, there is no direct expenditure of metabolic energy by the sodium co-transport process. The energy expenditure is indirect and results from the direct expenditure of energy by the Na,,K"-ATPase pump in creating and maintaining the sodium electrochemical gradient. This is the definition of secondary active transport, with glucose transport being secondary to the active transport of sodium. Many organic nutrients, including glucose, amino acids, several vitamins, and bile acids, are absorbed by sodium co-transport processes.
In addition to sodium co-transport, there are other types of secondary active transport pathways. These pathway proteins are known as exchangers or antiports. Exchangers are usually involved with ion transport and are similar to cotransport proteins in that they have binding sites for selected ions. The difference between exchangers and co-transport proteins is that for exchangers, the binding sites for the two different ligands are on opposite sides of the plasma membrane. For example, an important exchanger is the sodium/ hydrogen (Na1ZH+) exchanger in the apical membrane. The protein has a binding site for Na+ and another for H+. When the sites are unoccupied, the Na1 site faces the intestinal lumen, and the H' site faces the interior of the enterocyte. When both sites are occupied, the protein flips, transporting H' out of and Na' into the cell, thus explaining the name exchanger, with H’ exchanged for Na,. As with co-transport, the force driving the exchange is the Na* electrochemical gradient across the cell membrane.
Another form of active transport, tertiary active transport, occurs via transport pathway proteins and is driven by electrochemical gradients that are established by secondary active transport. The best example of tertiary active transport is the chloride/bicarbonate (C17HCO√) exchanger. This mechanism occurs in response to gradients established by the Na1ZH* exchanger, a secondary active transport mechanism. The Cl ZHGOf exchanger is discussed in more detail later, in the section on absorption. In essence, the term tertiary is used because the Na1jK+-ATPase system (primary) establishes the gradient that drives the Na7H~ exchanger (secondary), which then establishes the gradient that drives the C17HCO3" exchanger (tertiary).