Skeletal System
This is a system consisting of individual organs often referred to as “bones. ” When these organs are viewed individually, it is often difficult to appreciate the interrelationships of their structure and function.
Let us Considersomeof thosefunctions. Articulationof Iheindividualbonesororgansof thissystemplaysa central role in providing the supporting mechanism and shape of the animal. Without a skeletal system, a mammal would be shapeless. Perhaps we would resemble an earthworm with hair and mammary glands.Body locomotion is another significant function of the skeleton. Bones, however, cannot move by themselves but serve as levers for lever systems producing what we commonly perceive as movement. The movement is produced by muscles that are attached to the bony levers whose fulcrums are the joints of the body [Figure 1 —1].
Although there are various classifications of joints, a junction between two bones, the system used here is that based upon the functional nature of the joint. In this system, a synarthrosis is a joint in which there is little or no movement, e.g., sutures between the bones of the skull. An amphiarthrosis, on the other hand, permits slight movement, e.g., between individual vertebrae in the vertebral column. Free movement between bones occurs around joints called diarthroses, e.g., between the head of the humerus or upper arm bone and the glenoid cavity of the scapula or shoulder blade.
Certain bony complexes provide protection for some of the more vulnerable soft and vital organs: the cranium protects the brain, the vertebral column protects the spinal cord, and the rib cage and sternum shield the heart, lungs, and major blood vessels. In addition, bones serve several other functions such as sites for blood cell formation or hemopoiesis, fat deposition, and mineral deposits of essential ions such as calcium and phosphate.
Contrary to what you may conclude after studying this system, bones are living organs. The skeletal systems of all vertebrates are constantly subjected to remodeling activities, during which, part of a bone may be entirely remodeled in a very short period of time, e. g., in humans the distal epiphysis of the femur is replaced approximately every six months. Since bony tissue is a reservoir for minerals such as calcium and phosphates, it is normal for
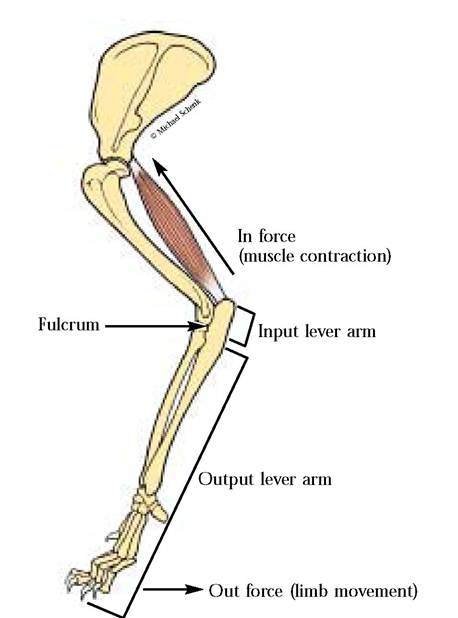
FIGURE 1-1 Muscles (Triceps brachii) and bones as lever systems.
mineral recycling to occur between bones and the blood. Calcium is essential for such physiological activities as muscle contraction, coagulation of blood, and the dynamics of membrane permeability, while phosphates are necessary for synthesis of nucleic acids, production of ATP, and metabolic events involving activation or deactivation of enzymes. Injuries to this living bony tissue require active repair processes.
During the study of the skeletal system, you will become familiar with distinctive processes and their positions, markings, foramina, and other features specific for each bone. These will allow you to distinguish left and right members of paired bones. It is important to identify the position of the bone in the articulated skeleton in order to appreciate the relationship of regional skeletal elements.
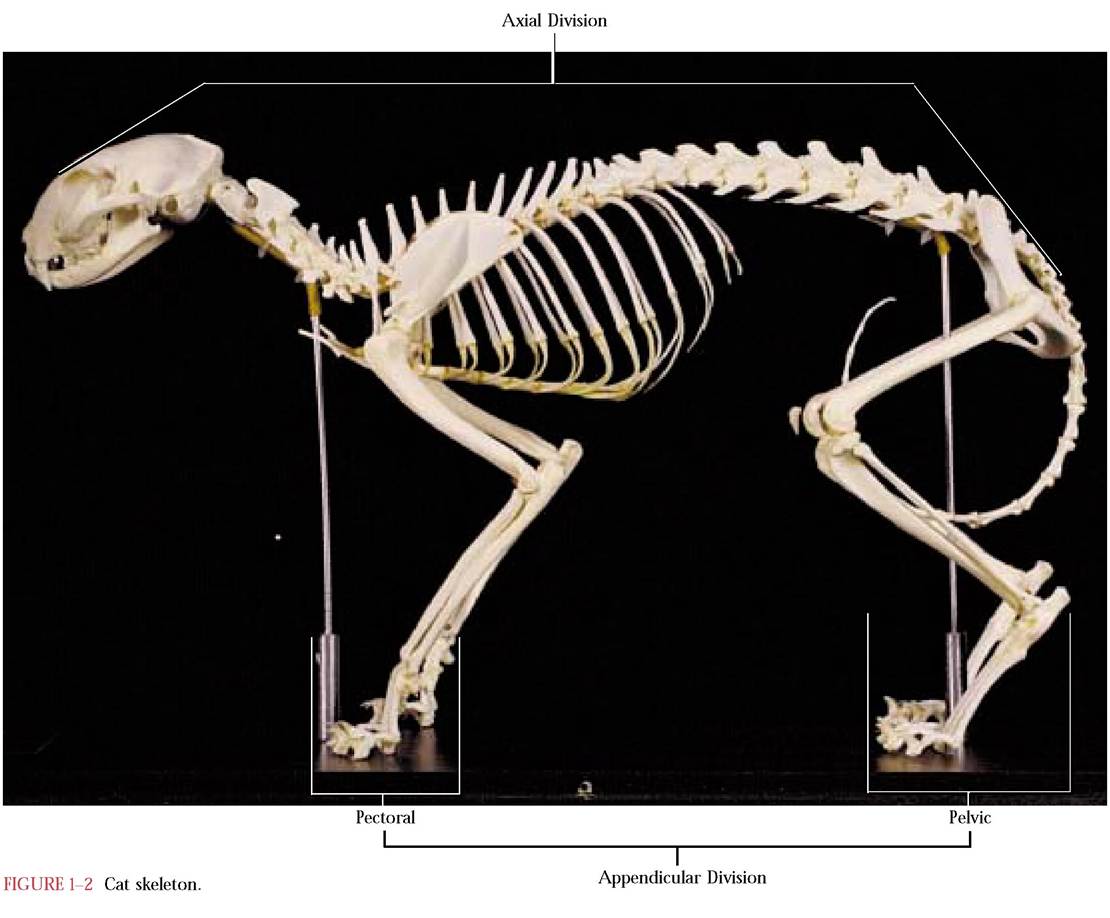
The skeleton consists of two parts known as the axial and appendicular divisions. The functions of these subdivisions are really very different from each other. The components of the axial division surround and protect soft body tissues and include the bones of the skull, the mandible, the hyoid, the vertebral column, the ribs, and the sternum. In contrast, the appendicular division is surrounded by soft tissues and can be further subdivided into the pectoral girdle with its forelimb and the pelvic girdle with its hindlimb [Figure 1-2].
AXIAL DIVISION
The Skull
The skull consists of the cranium whose bones surround the brain and permit the cranial nerves to enter or exit through a number of openings known as foramina, fissures, and
canals.* Closely associated with the cranium are a number of facial bones that give the face its shape. This essential complex (cranial and facial bones) contains a number of cavities that surround and protect the major sense organs such as the eyes, portions of the auditory apparatus, olfactory organs, and with the articulated mandible, the gustatory organs, all of which have evolved as specialized organs closely associated with the central nervous system [Figure 1-3A, Figure 1-3B, and Figure 1-3C].
As you examine the individual bones of the skull, refer to the above figures for positions of the bones relative to one another.
The Premaxilla
The paired palatine processes of the premaxillae [Figure 1-3C and Figure 1-4B] occurring at the most anterior margin of the upper jaw form approximately one-sixth of the hard palate. In the cat, three incisors are rooted in each premaxilla. However, only two incisors are found in humans, one central and one lateral. Additionally, the premaxilla is fused with the maxilla and is not identifiable as a separate bone. The incisors are located in the central portion of the human maxilla in the position where the premaxilla can be identified in many other mammals. A pair of openings, the anterior palatine foramina (incisive ducts), can be seen just posterior to the incisors in the ventral view. In the human a single incisive foramen occurs in a comparable position in the maxilla. The premaxillae are bordered by the nasal bones dorsally, and laterally and caudally by the maxillae. With the nasals the premaxillae form the anterior edge of the nasal cavity.
*In the discussion of the skull, there will be numerous references to branches of cranial nerves (C.N.) as follows:
C.N.
I—OlfactoryC.N. II—Optic
C.N. III—Oculomotor
C.N. IV—Trochlear
C.N. V—Trigeminal
C.N. VI—Abducens
C.N. VII—Facial
C.N. VIII—Vestibulocochlear
C.N. IX—Glossopharyngeal
C.N. X—Vagus
C.N. XI—Spinal Accessory
C.N. XII—Hypoglossal
In the text, reference will be made in the following manner, e.g., C.N. V, C.N. XII, etc.
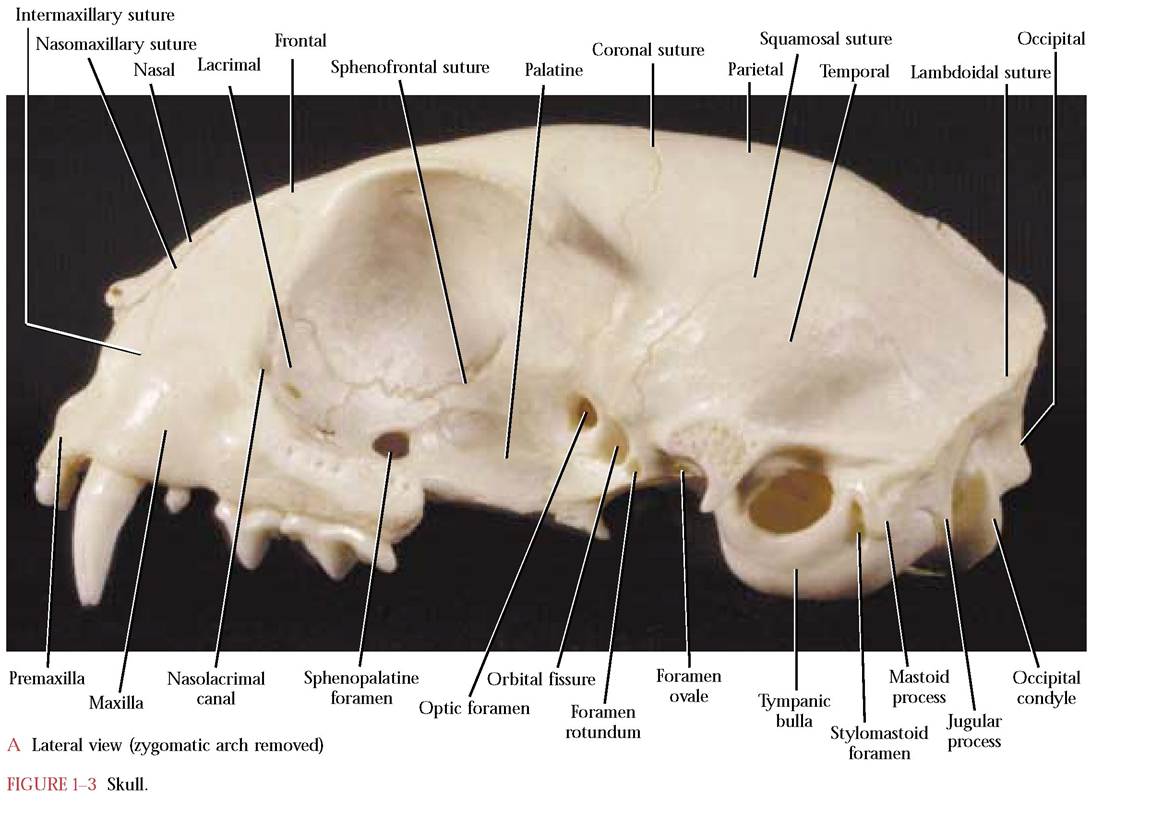
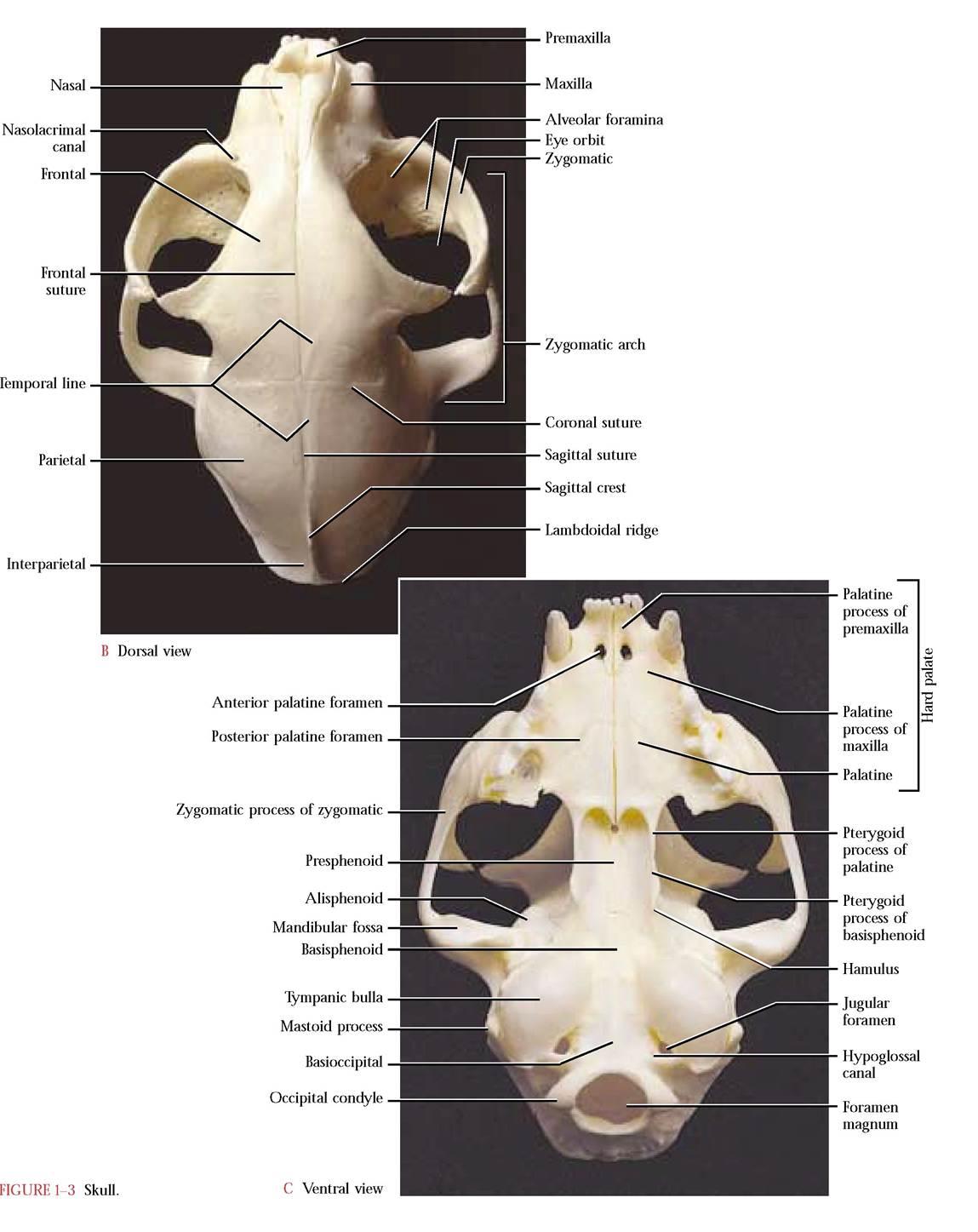
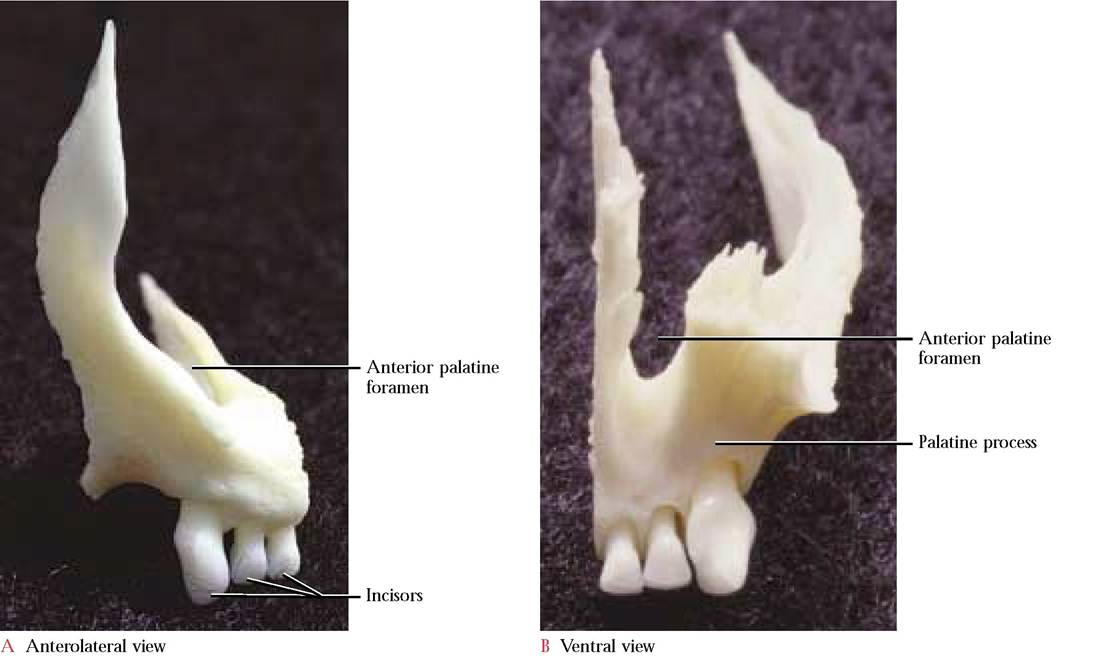
FIGURE 1-4 Right premaxilla.
The Maxilla
Lateral to the premaxillae are the paired maxillae [Figure 1-5a and Figure 1-5B] that complete the upper jaw and contribute to the hard palate ventrally as shelves of bone known as the palatine processes of the maxillae. These processes extend back to the posterior border of the hard palate laterally but are joined medially by paired palatine bones to complete the complex that we call the hard palate [Figure 1-3C]. In contrast, the contribution by the palatine bones in the human is greatly reduced. In the cat, each maxilla articulates with the zygomatic or malar, the premaxilla, nasal, lacrimal, frontal, and palatine bones and extends into the eye orbit as the anterior floor.
At the anterior border of the maxilla is a perceptible bulge in the alveolar process that accommodates the extremely long root of the canine tooth. Posterior to the canine is the smallest of three premolar teeth. Posteromedial to the third premolar is a vestigial molar. The roots of all of these teeth are embedded within the alveolar process of the maxilla.
Dorsal to the canine and first premolar tooth is the elongated, flat, thin frontal process that is wing-shaped.
The infraorbital foramen pierces the maxilla at the junction of the body and frontal process. Through this foramen pass the infraorbital blood vessels and infraorbital nerve (C.N.
V). Take note of the many small openings, the alveolar foramina, in the floor of the orbit through which small branches of the infraorbital nerve (C.N. V) pass to the teeth in the upper jaw [Figure 1-3B]. At the posterior end of the body is the zygomatic process. On the medial aspect of the canine alveolar elevation exists a small lateral contribution to the nasal conchae, the maxilloturbinate. In humans, it appears that the maxilloturbinate is a separate bone and is identified as the inferior nasal concha. No maxillary sinuses occur in the cat, however, each human maxilla contains one.The Nasal
Medial to the premaxilla and maxilla and cranial to the frontals are the paired nasal [Figure 1-6] bones that form the dorsal wall of the nasal cavity. Another small dorsal contribution to the nasal conchae, the nasoturbinate, is present on the ventromedial aspect of the nasals. Humans do not have a homologous contribution to the turbinates from the nasals.
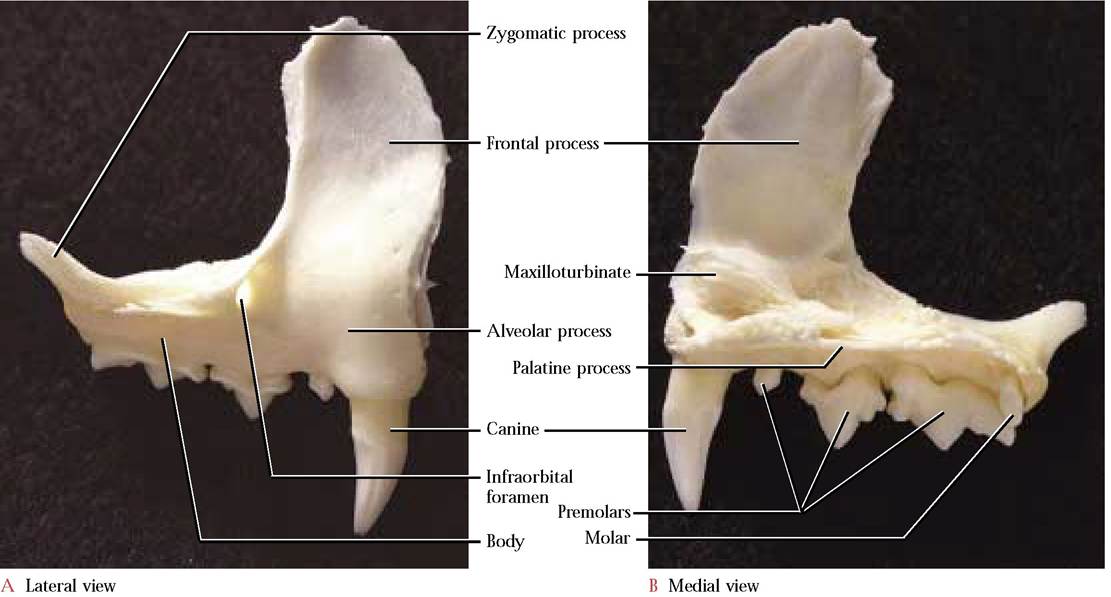
FIGURE 1-5 Right maxilla.
Nasoturbinate
FIGURE 1-6 Nasal: right cranial, left caudal views.
The Ethmoid
The ethmoid, an unpaired cranial bone [Figure 1-7A and 1-7B], is entirely associated with the nasal cavity. The major bulk of the ethmoid, the ethmoturbinates, composed of elaborately scrolled, thin laminate plates of bone, comprise most of the nasal conchae and fill most of the nasal cavity. In humans, the superior and middle nasal conchae are homologous with the ethmoturbinates in the cat. The thin perpendicular plate of the ethmoid separates the two lateral ethmoturbinates and with the vomer forms the bony portion of the nasal septum. Caudal to the perpendicular plate is the thin, concave, perforated cribriform plate. Through these foramina pass fibers of the Olfactory Nerve (C.N. I) that synapse with neurons that lie in the olfactory bulbs situated directly above the cribriform plate.
In humans, a triangular, dorsal projection from the cribriform plate, the crista galli, serves as an anterior point of attachment of the meninges (protective brain membranes). Additionally, in contrast to the cat, there are extensive sinuses in the ethmoid of humans.The Vomer
Ventral to the ethmoid and dorsal to the hard palate is the single vomer [Figure 1-8] that completes the bony nasal septum. The dorsal, V-shaped, groove of the vomer cradles the perpendicular plate of the ethmoid while the lateral
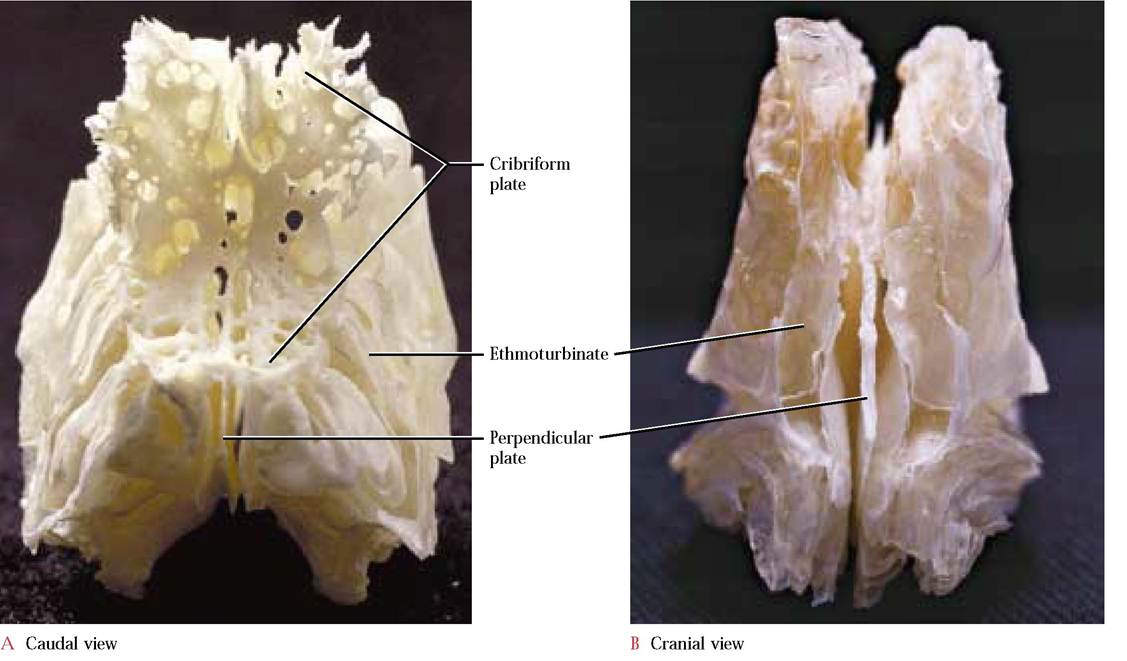
FIGURE 1-7 Ethmoid.
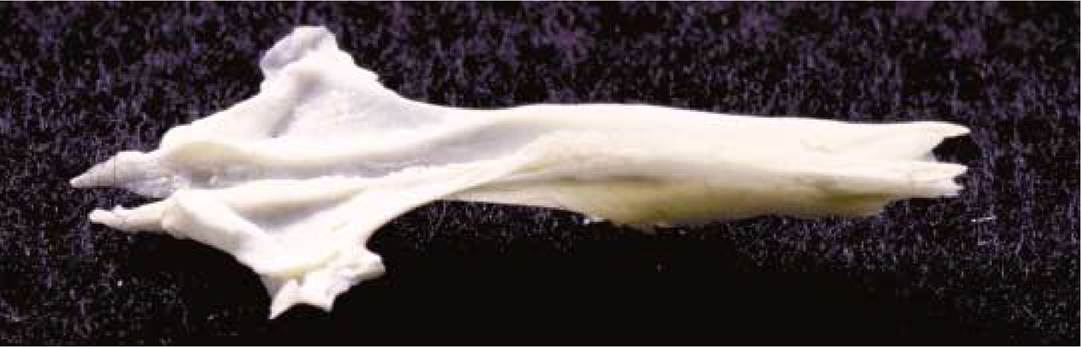
FIGURE 1-8 Vomer: dorsal view.
wings articulate with the ethmoturbinates. Its cranial end articulates with the palatal processes of the premaxilla, the ventral region with the palatine processes of the maxilla while the caudal end articulates with the presphenoid.
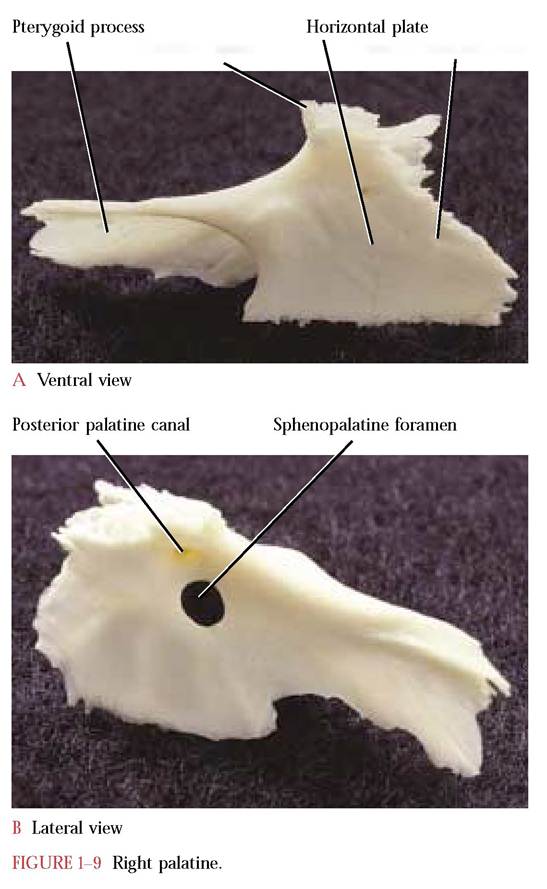
The Palatine
Posterior to the palatine processes of the maxillae are the paired palatine bones [Figure 1-9A and Figure 1-9B]. The palatine bone consists of a horizontal plate and a curved irregularly shaped vertical plate. The paired horizontal plates of the palatines meet in the mid-ventral line and form the caudal end of the hard palate. Two small foramina, the cranial openings of the posterior palatine canals through which the greater palatine nerve of the maxillary division of the Trigeminal Nerve (C.N. V) pass, are found within the lateral portion of the horizontal plates. The entire
Maxillary spine
Posterior palatine canal
antero-lateral surface of these plates articulates with the palatine processes of the maxilla and terminates posteriorly with the maxillary spine. The narrow postero-lateral extensions of the vertical plates form the pterygoid processes of the palatine that articulate with the pterygoid processes of the basisphenoid. These palatine processes continue dorso- medially as thin, curved, laminar plates that articulate with the presphenoid. The pterygoid processes in the human have been abbreviated into the vertical palatine plates that are intimately associated with the greater wing of the sphenoid. The larger anterior third of these vertical plates is pierced by the sphenopalatine foramen carrying the sphenopalatine nerve (C.N. V) and corresponding blood vessels. A smaller opening, the posterior palatine canal, occurs slightly cranial and lateral to the sphenopalatine foramen. Through it passes the greater palatine nerve of the maxillary division of the Trigeminal Nerve (C.N. V) and corresponding artery. These plates articulate dorsally with the frontal bone and cranially with the lacrimal bone.
The Lacrimal
The small wafer-like bone that is roughly shaped like a rhombus and located in the antero-medial portion of the eye orbit, is the lacrimal [Figure 1 —10]. The nasolacrimal canal is a prominent landmark of the anterior portion of the eye orbit. The canal includes an anterior notch in the lacrimal and is completed by a groove in the frontal process of the maxilla with which it articulates. In addition, the lacrimal articulates with the frontal and palatine.
The Frontal
The cranial portion of the skull roof consists of the paired frontals [Figure 1-11A and Figure 1-11B]. Dorsally, the
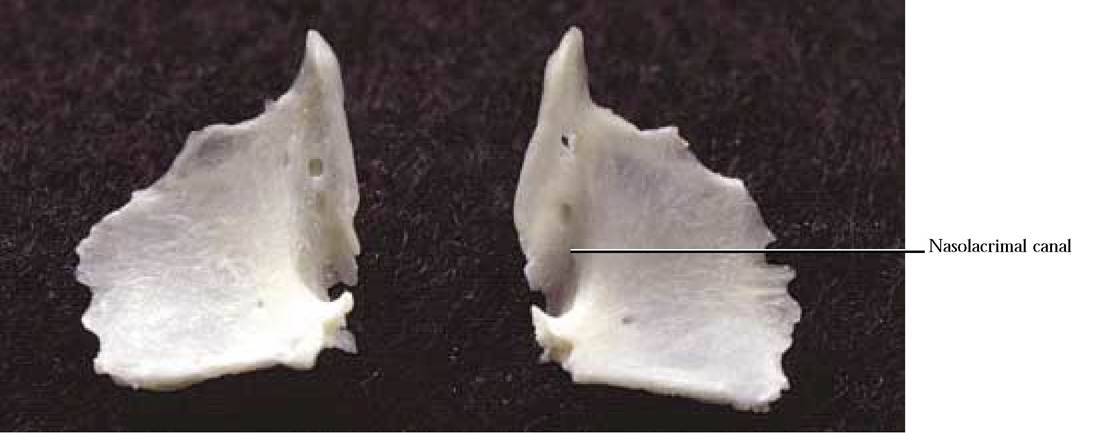
FIGURE 1-10 Right and left lacrimal: lateral view.
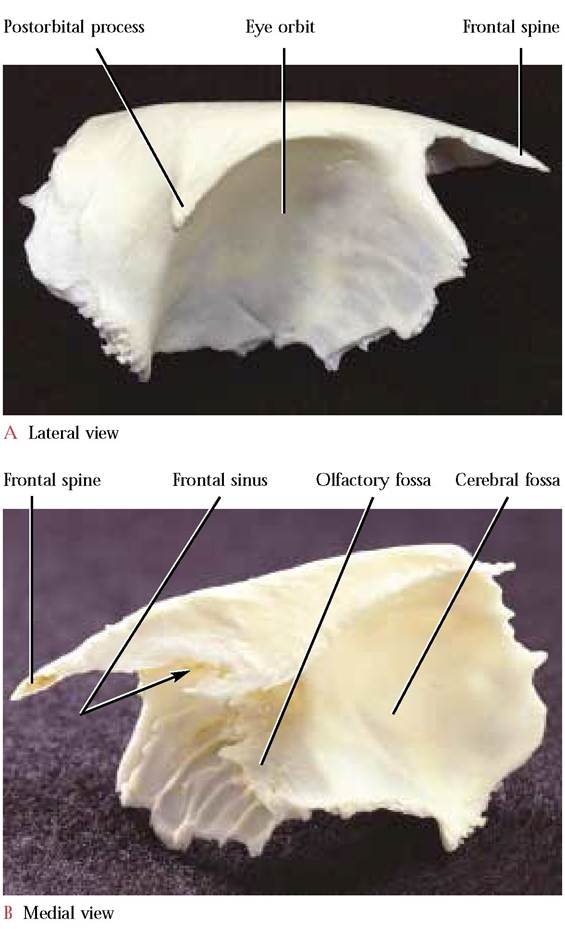
FIGURE 1-11 Right frontal.
articulated frontals form a smooth surface that resembles a short Chinese pagoda. The anterior frontal spines form the roof-peak of the pagoda that articulate with the nasals and maxillae while the postorbital processes complete the lower edge of the roof.
The caudal surface that articulates with the parietals, forms the lower part of the building. The lateral surface, prominently concave, shapes the dorsomedial and posterior aspect of the orbit. Articulation of the frontal with the lacrimal, ethmoid, maxilla, presphenoid, and palatine bones completes the medial and ventral part of the eye orbit.
The medial view reveals a number of distinctive features. The obvious vertical plate abuts its partner at the site of the external sagittal suture and together they join the perpendicular plate of the ethmoid located ventrally. By peering into the anterior ends of the articulated frontal bones one can observe a recessed oval foramen leading into the extensive frontal sinus. The furrowed ethmoid surface on the cranio-medial wall of the eye orbit mirrors the surface of the nasal conchae, since it is in this area that the ethmoid and the frontal closely approach one another. Along the posterior border of this area is a curved ridge, extending from the ventral edge of the frontal sinus opening to the ventral border of the frontal bone, that matches the curvature of the cribriform plate with which it articulates. A number of minute, inconspicuous foramina in the medial orbital surface of the frontal are often difficult to see. These openings allow the passage of small blood vessels and nerves. In the human, a supraorbital notch or foramen transmitting corresponding nerves and veins occurs along the supraorbital margin. Note the olfactory fossa which houses the olfactory lobes and the anterior portion of the cerebral fossa containing the anterior part of the cerebrum [Fig. 1-11B].
The Zygomatic
The gently curved surface of the zygomatic or malar bone [Figure 1-12] forms the lateral boundary of the eye orbit. Two projections occur posteriorly. A dorsal orbital process curves up toward the postorbital process of the frontal to surround the orbit with an incomplete bony ring. In the human, the bony ring is complete. The temporal fenestra (opening) inherited by mammals (e.g., the cat) from their “reptilian ” ancestors has been enclosed largely by the greater wing of the sphenoid. A ventral zygomatic process articulates with the zygomatic process of the temporal, forming the “cheek bone” or zygomatic arch. A distinct ridge along the lateral surface marks the position of the origin of the masseter muscle. The curved anterior end is irregularly serrated and fits into a trough in the maxilla.
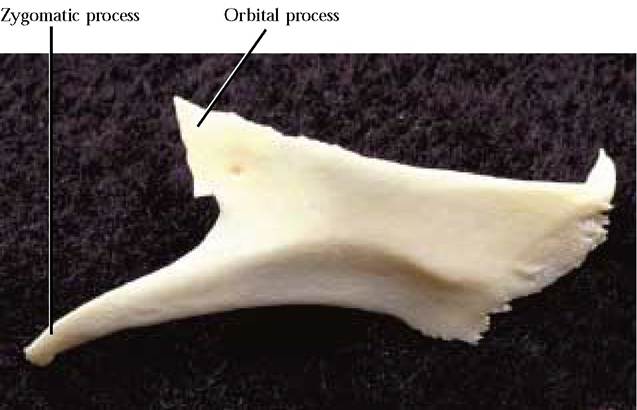
FIGURE 1-12 Right zygomatic: lateral view.
The Parietal
Caudal to the frontal and forming the posterior portion of the cranial vault are the paired parietals [Figure 1-13A, Figure 1-13B, and Figure 1-14]. Each bone is roughly rectangular in shape with a smooth, curved outer surface. The articulation between the paired parietals completes the
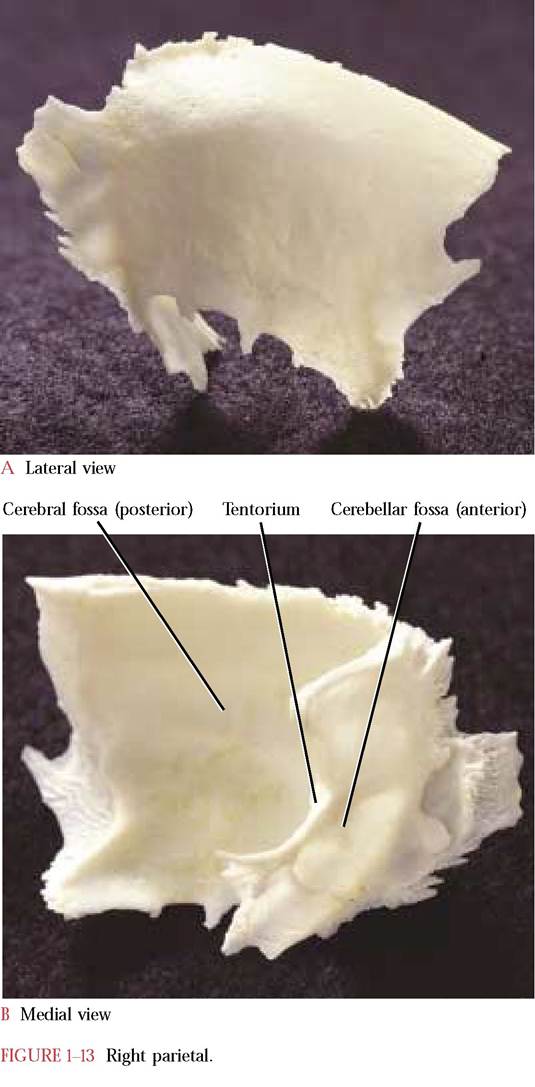
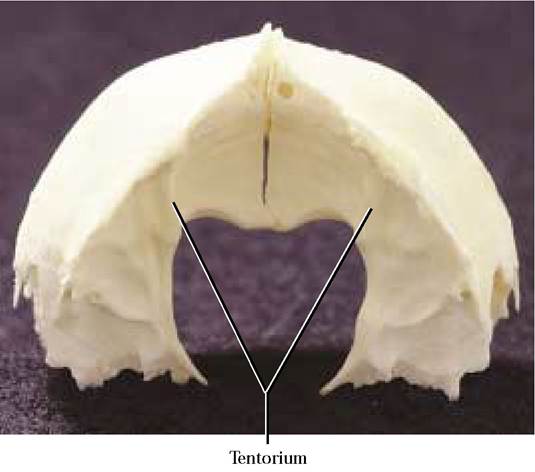
FIGURE 1-14 Paired parietals: caudal view.
sagittal suture (also described in the discussion of the frontal bone). The parietals at their caudal ends diverge to accommodate the articulation with the dorso-medially located interparietal (in a disarticulated skull the interparietal may be a separate bone or may remain attached to one of the parietals). The occipital articulates with the parietal- interparietal complex. The lateral edge articulates with the squamosal portion of the temporal.
The concave anterior two-thirds of the medial surface reflects the gyri (elevations) and sulci (furrows) of the parietal lobe of the cerebral hemisphere of the brain and constitutes the posterior portion of the cerebral fossa. The tentorium projects anteriorly as a curved shelf with a prominent medial notch that together with its partner forms a large opening that is shaped like the upper part of a valentine. Through this large foramen passes the brainstem of the central nervous system. Caudal to the tentorium is the anterior portion of the cerebellar fossa. The irregular surface of this fossa also reflects the gyri and sulci of the cerebellum.
The Interparietal
A single, triangular bone located between the parietals and the occipital is the interparietal [Figure 1-15]. The dorsal surface is distinguished by a medial projection called the sagittal crest that in some cats extends anteriorly onto the surface of the parietals. Posteriorly, this crest is continuous with the lambdoidal crest of the occipital. The interparietal is quite conspicuous in young cats, however, in older cats is generally fused with the parietals. In the human, the interparietal may have been lost or at least cannot be distinguished.
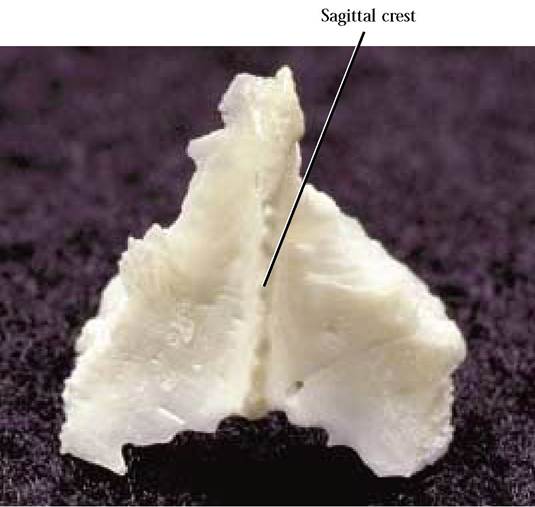
FIGURE 1-15 Interparietal: caudal view.
The Occipital
The occipital bone [Figure 1-16A and Figure 1-16B] completes the posterior portion and base of the skull. From the posterior aspect, this skull bone resembles an equilateral triangle that has curved sides, breaking the rules of geometry. A large opening, the foramen magnum, is situated in the ventral part of the occipital. The foramen magnum marks the site of transition of the brain into the spinal cord as that portion of the central nervous system enters the vertebral canal.
The basal part, the basioccipital, is flat and elongated, and articulates with the basisphenoid portion of the sphenoid anteriorly and the petrous part of the temporal bone laterally. The lateral portions, the exoccipitals, articulate with the petrous and mastoid portions of temporal. Notice the prominent occipital condyles with which the atlas or first cervical vertebra articulates. Lateral to each condyle and separated by a deep depression is a short, blunt projection, the jugular process, at the vertices of the triangle base. This projection overlaps the tympanic bulla of the temporal, with which it articulates. At the site of this articulation is an opening, the jugular foramen, through which pass the Glossopharyngeal (C.N. IX), the Vagus (C.N. X) and the Spinal Accessory (C.N. XI) cranial nerves along with the internal jugular vein. Medial to the jugular foramen and located in a notch associated with the wall of the foramen is the hypoglossal canal through which the Hypoglossal Nerve (C.N. XII) passes. As one looks through the foramen magnum, the extremely variable condyloid canal, through which passes a small vein or veins, can be identified dorsal to the hypoglossal canal [Fig. 1-21].
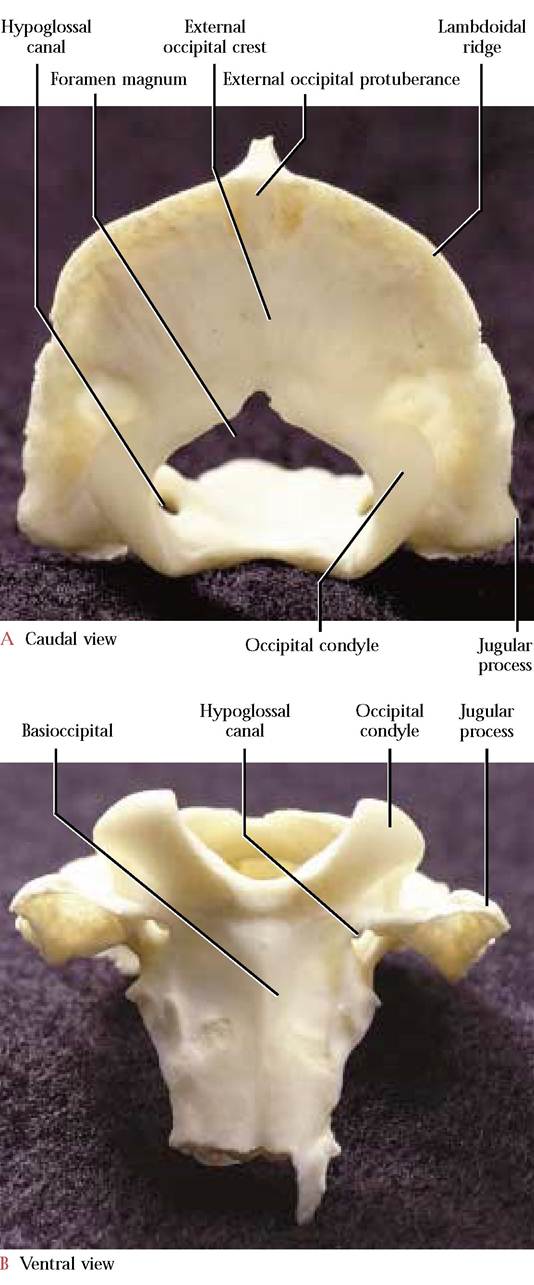
FIGURE 1-16 Occipital.
The Supraoccipital or squamous portion forms an arch over the foramen magnum and articulates with the parietals and interparietals. The sharp edge of the arch that extends from one jugular process to the other is known as the lambdoidal ridge (nuchal crest). An inconspicuous median crest, the external occipital crest, extends from the lambdoidal ridge to the upper rim of the foramen magnum. The external occipital protuberance is located at the junction of this crest with the lambdoidal ridge.
The Sphenoid
Anterior to the occipital sits the unpaired sphenoid [Figure 1-17A, Figure 1-17B, and Figure 1-17C] whose shape has often been described as resembling a butterfly. The body of the “butterfly ” is known as the basisphenoid whereas the wings are referred to as the alisphenoids. Two anteriorly oriented projections termed the pterygoid processes have fused with the alisphenoids. In mammalian ancestors these processes were separate pterygoid bones. The hamulus, a thin rod, extends posteriorly from the body of the pterygoid process. As previously mentioned, these processes articulate with the pterygoid processes of the palatine. Dorsal to the pterygoid process are three foramina. The cranial margin of the first of these openings, the orbital fissure, is incomplete, however, the articulation of the presphenoid with the sphenoid at this site completes the cranial margin of the fissure [Figure 1-18]. Through this opening passes the Oculomotor Nerve (C.N. III), the Trochlear Nerve (C.N. IV), the Ophthalmic division of the Trigeminal Nerve (C.N. V) and the Abducens Nerve (C.N. VI). The single orbital fissure of the cat is represented in the human by a superior orbital fissure that is continuous with an inferior orbital fissure located in a comparable position. Like the orbital fissure in the cat, similar nerves and blood vessels are transmitted through these openings. The middle opening of this trio of foramina in the cat is the foramen rotundum, through which passes the Maxillary branch of the Trigeminal Nerve (C.N. V). The third opening is the foramen ovale, through which exits the Mandibular division of the Trigeminal Nerve (C.N. V). In the human there is an additional small foramen, the foramen spinosum, that is located postero-lateral to the foramen ovale. Through this foramen passes a branch of the mandibular nerve and blood vessels. Laterally, the dorsally curved portion of the
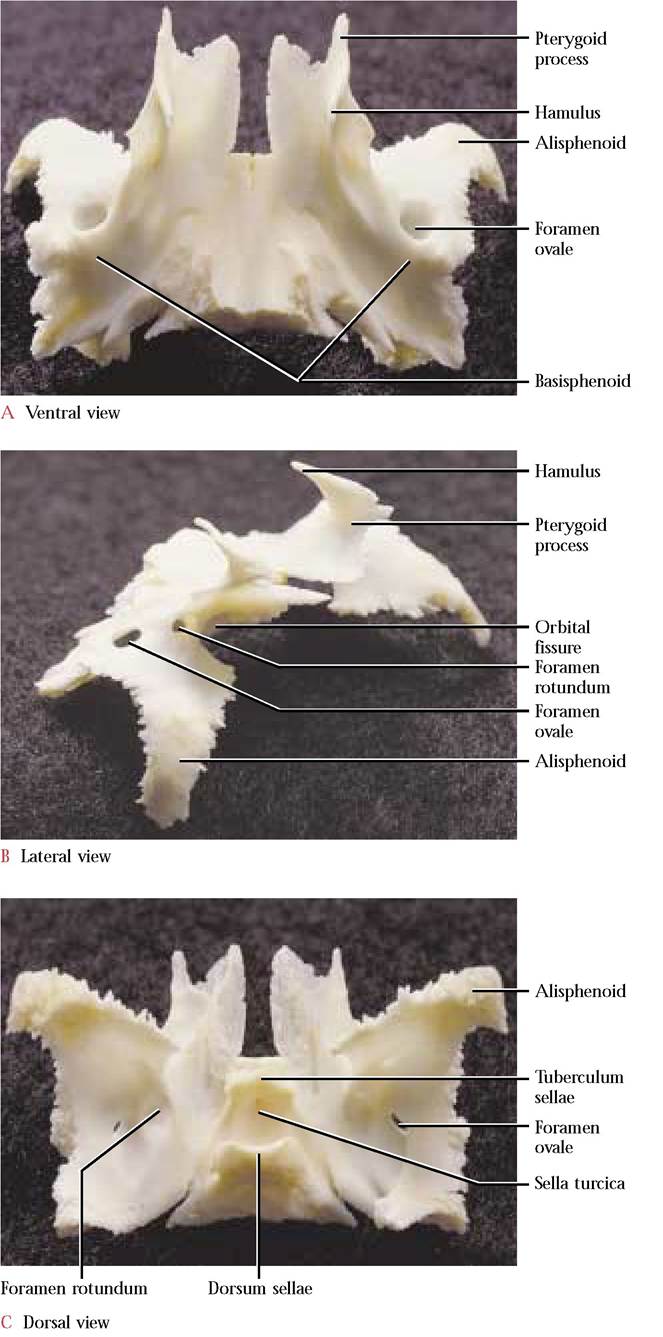
FIGURE 1-17 Sphenoid.
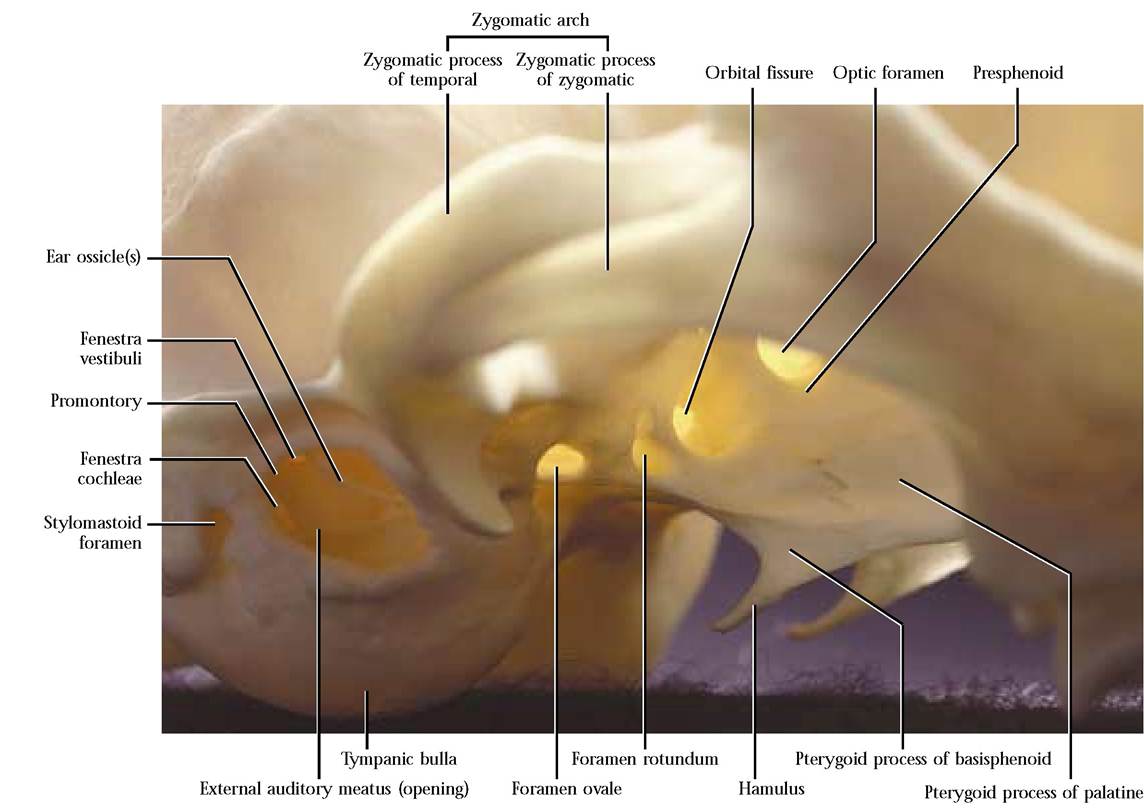
FIGURE 1-18 Sphenoid, presphenoid, tymbanic bulla region (enlarged).
“butterfly” wing, the alisphenoid, articulates with the squamous portion of the temporal, while the caudal portion of the wing articulates with the tympanic bulla.
Ventrally, the body of the basisphenoid is rather smooth and has a slight medial ridge, a continuation of the ridge present on the ventral surface of the basioccipital with which it articulates, posteriorly. The anterior end of the basisphenoid articulates with the body of the presphenoid. The dorsal or inner surface of the basisphenoid is distinctly saddle-shaped. The anterior elevation of the saddle is the tuberculum sellae whereas the more prominent and rounded posterior elevation is the dorsum sellae. Between the two elevations is a conspicuous depression known as the sella turcica in which the hypophysis (pituitary gland) rests. A nutrient foramen occurs posterior to the tuberculum sellae and a pair of similar foramina are present posterior to the dorsum sellae.
The Presphenoid
The unpaired presphenoid [Figure 1-19A and Figure 1—19B] consists of a body and two wings. The ventral surface of the body resembles an hourglass with a deep anterior notch and a rounded posterior base. A median ridge, fairly prominent posteriorly, is continuous with the ridge of the basisphenoid. Two articulating facets lateral to the ridge accommodate the pterygoid processes of the sphenoid. As one peers into the notched anterior end of this bone two conspicuous sphenoidal sinuses, separated by a median longitudinal bony partition, are evident. These sinuses end blindly at the posterior end of the body of the presphenoid. The wings project postero-laterally and resemble small triangles. Lateral to the body and piercing the base of the triangle is the optic foramen through which the Optic Nerve (C.N. II) and ophthalmic artery pass. On the dorsal or inner aspect, the presphenoid is smooth and distinguished posteriorly by the transverse chiasmatic groove that extends between the optic foramina. The chiasmatic groove is the site of the optic chiasma. The human sphenoid consists of a complex resulting from the fusion of the presphenoid and sphenoid and although the individual bones are not distinct, the structures, processes, and foramina identifiable in the cat are found in comparable positions.
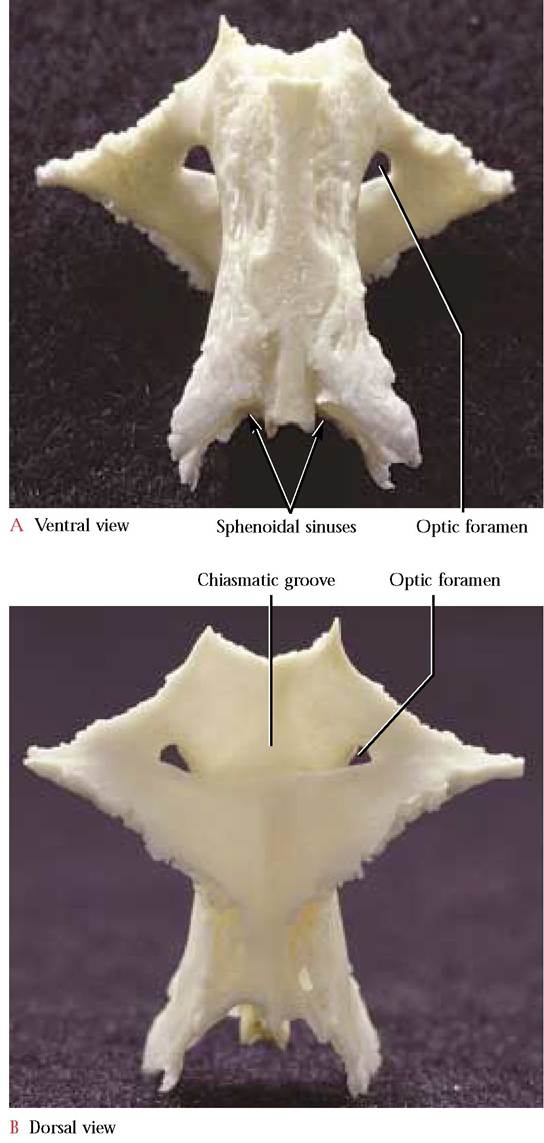
FIGURE 1-19 Presphenoid.
The Temporal
When turned upside down, the temporal bone [Figure 1-20A and Figure 1-20B] resembles a young nestling bird stretching its wing. This is a complex bone that consists of the squamous, the petrous, and the tympanic portions. The squamous part represents the breast of the “nestling, ” and is thin and smooth laterally while the medial surface reflects the contours of the cerebellum that rest against it. The zygomatic process, a distinctly curved, anteriorly oriented projection representing the “wing” of the bird, extends from its ventrolateral surface. The anterior half of this process is beveled along the ventral border to accomodate the zygomatic process of the zygomatic (malar) bone. A groove, the mandibular fossa, in the posterior part of the process provides the articulating surface for the condyloid process of the mandible. The small postmandibular process, projecting from the proximal basal portion of the zygomatic process, functions as a posterior brace for the condyloid process of the mandible.
The tympanic portion (head of the bird) is roughly ovoid and hollow. This portion is the tympanic or auditory bulla [Figure 1-18]. An irregular oval opening on the lateral surface, the external auditory meatus (eye of the bird), leads into the tympanic cavity. Often the malleus and incus, two of the three middle ear ossicles can be identified within the tympanic cavity. The third ossicle or stapes is not visible since it is not only small, but is oriented more medially and fits into the oval window of the inner ear. However, if the ossicles are not present, it is possible to see several openings, the fenestra cochleae (round window), the fenestra vestibuli (oval window), and the foramen leading to the facial canal. The opening of the round window is located below and passes under an elevated rim, the promontory, in a fashion similar to a tunnel entering a mountain. The oval window is located dorsal to the promontory and is oriented at approximately a right angle to the tunnel mentioned above. Caudal to the fenestra ovalis and dorsal to the tunnel rim is a small groove leading into the facial canal permitting passage of the Facial Nerve (C. N. VII) through the petrous portion of the temporal. Notice the distinct thin ridge in the floor of the auditory canal extending along a more-or-less anterior to posterior line. This is the dorsal edge of the tympanic bulla where it articulates with the petrous portion of the temporal.
The mastoid portion of the petrous region (beak of the bird) is elongated posteriorly and articulates with the lateral portion of the occipital. A prominent, angular, nipple-shaped projection, the mastoid process, overlaps the bulla posterior to the external auditory meatus. The stylomastoid foramen, occurring anterior to the mastoid process permits passage of the Facial Nerve (C.N. VII). A distinct opening, the internal auditory meatus, occurs in the medial surface. A bony partition divides the meatus into a dorsal canal through which passes the Facial Nerve (C.N. VII) and several ventral foramina for the passage of the Vestibulocochlear Nerve (C.N. VIII). Within the petrous bone the membranous acoustic and equilibrium apparatus resides in the bony labyrinth that is slightly larger but of a similar shape. Dorsal to the meatus is a small, deep fossa, the appendicular fossa, that reflects the position of the small appendicular lobe of the cerebellum. A conspicuous elongated slit between the petrous portion and the tympanic bulla admits the eustachian tube into the middle ear cavity.
The temporal bone in the human, with some minor modifications, is quite similar to the cat. Perhaps one of the most obvious differences is the absence of a tympanic bulla,
however, the tympanic or middle ear cavity of the bulla that houses the ear ossicles is certainly present and the outer of the temporalis muscle in individual cats. Older cats often have more prominent temporal lines. The extent of the sagittal crest also varies with the age of the cat. In older, more mature animals, it extends from the lambdoidal ridge onto the posterior surface of the parietals. Its height and rugosity, likewise, varies with age.
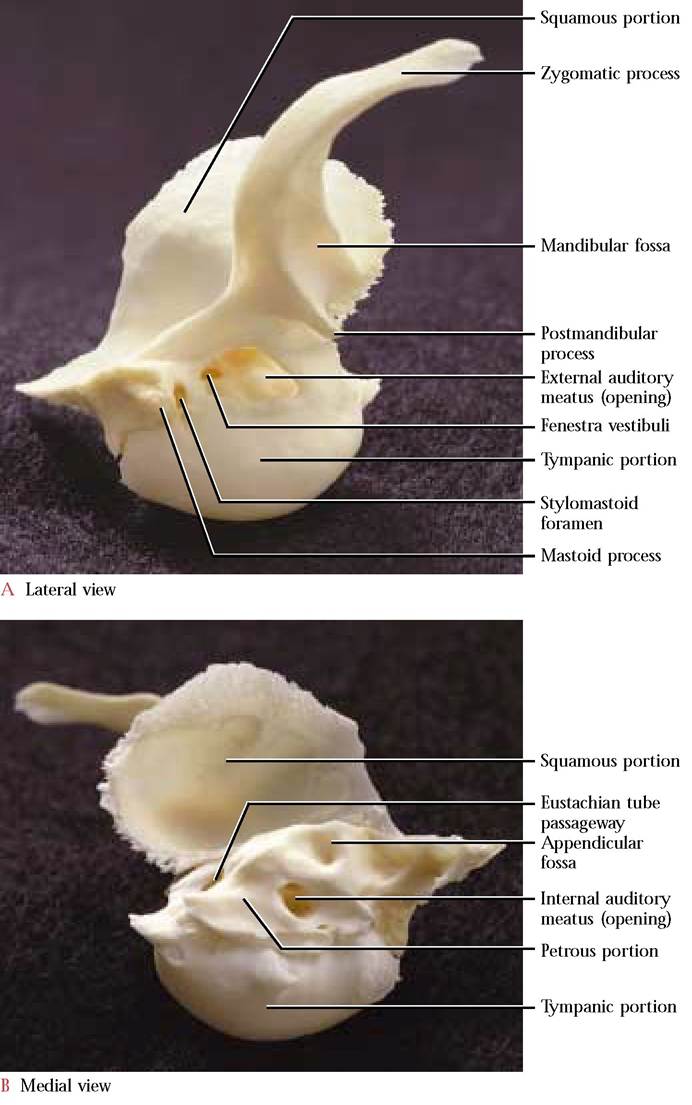
FIGURE 1-20 Right temporal.
covering of the region is represented by the much elaborated mastoid process that in the cat could be overlooked by the careless student. A pointed, downward directed styloid process, not present in cats, is evident just anterior to the stylomastoid foramen.
Surface Features
Although the overall skull topography is fairly smooth, there are some conspicuous features such as sutures, lines and ridges, and openings [Figure 1-3A, Figure 1-3B, and Figure 1-3C]. Sutures, synarthrotic joints, are immovable articulations found between adjacent skull bones and are generally identified using the names of the bones between which they occur.
Intermaxillary: between the premaxillary and maxillary bones
Nasomaxillary: between the nasal and maxillary bones
Sphenofrontal: between the sphenoid and frontal bones
Frontal: between the frontal bones Coronal: between the parietal and frontal bones
Sagittal: between the parietal bones Squamosal: between the parietal and squamous portion of the temporal bone Lambdoidal: caudad of the parietals, separating them from the occipital and interparietal
A faint dorsal feature is the temporal line that demarcates the origin of the temporalis muscle. It extends from the caudal margin of the postorbital process as a gentle curve over the frontal bone onto the parietal bone meeting its opposite partner in a V-shaped pattern terminating at the cranial end of the sagittal crest. Some variability in the extent of the temporal line is seen dependent upon the age-related development
Note the prominent, lateral curved bar or “cheekbone, ” the zygomotic arch, composed of a posterior zygomatic process of the temporal that overlaps an anterior zygomatic process of the malar or zygomatic.
Cavities and Sinuses
Mammal skulls possess a number of spaces, cavities, and sinuses which reduce the weight of the skull and protect and house sense organs [Figure 1—21]. The nasal cavity, the most anterior of these spaces, houses the highly convoluted turbinates of the ethmoid, maxilla, and nasal bones whose mucous membrane plays an essential role in humidifying, warming, and filtering of respiratory air. It is also within this membrane that olfactory receptors associated with the sense of smell occur. A nasal septum, consisting of the dorsal bony perpendicular plate of the ethmoid, a rostral cartilaginous portion, and the ventrally located vomer, divides the nasal cavity into right and left halves. The external nares (nostrils) open anteriorly into these cavities. The caudal cribriform plate of the ethmoid through which pass the olfactory fibers, separates the nasal cavity from the cranial cavity.
The cranial cavity is organized into four distinct areas. The olfactory fossa occurs just caudal to the cribriform plate and houses the olfactory bulbs where the olfactory fibers synapse. Posterior and continuous with the olfactory fossa is the cerebral fossa that houses the cerebrum, the diencephalon, and the mesencephalon (midbrain). In the floor of the cranial cavity is the sella turcica in which the hypophysis (pituitary) sits. The tentorium marks the caudal end of the cerebral fossa. Posterior to the tentorium lies the cerebellar fossa which surrounds the pons, cerebellum, and the medulla oblongata. A small deep depression, the appendicular fossa, lies dorsal to the internal auditory meatus in the petrous portion of the temporal bone. The foramen magnum opens at the caudal end of the cerebellar fossa. Notice the considerable topography of the surface of the cranial cavities which mirrors the contours of the external brain surface and also the blood vessels associated with the brain.
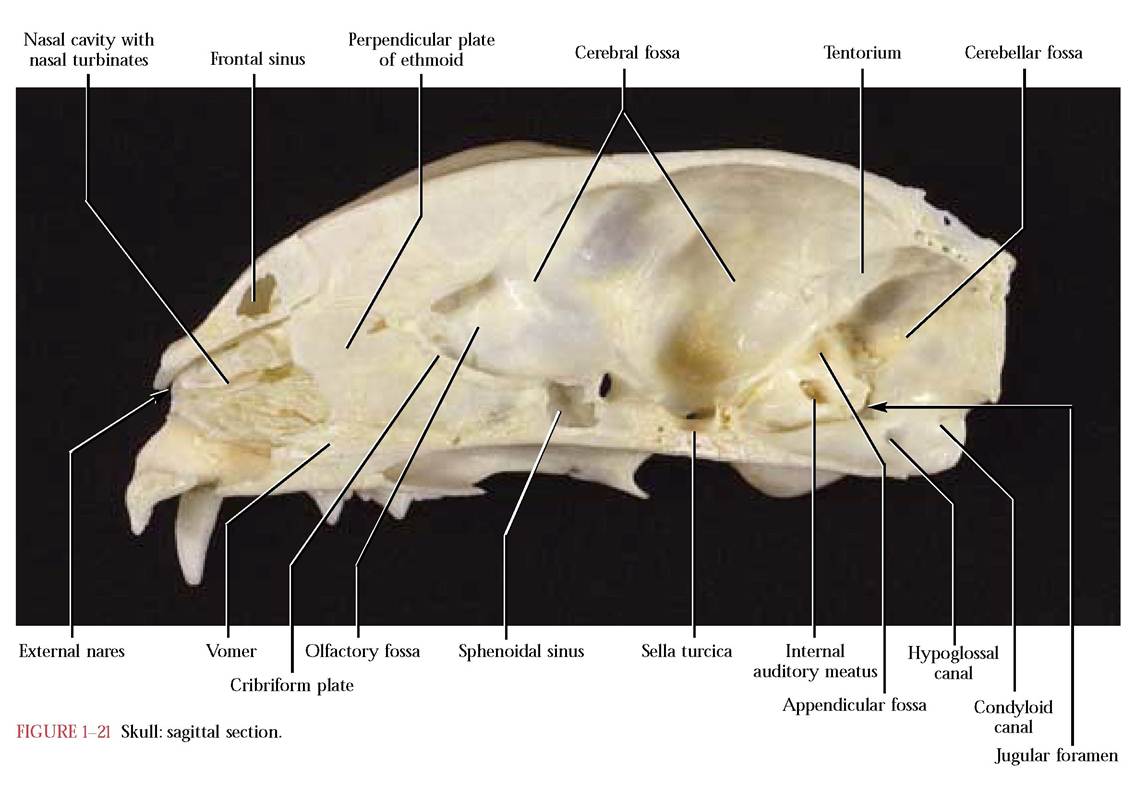
Two air-filled spaces, the frontal sinus in the frontal bone and the sphenoidal sinus enclosed in the presphenoid bone of the skull probably function in weight reduction and in many mammals, e.g., the cat, the olfactory epithelium extends into the sinuses, thereby enhancing the sense of smell. Consequently, some mammals smell better than humans. In addition, in the human, sinuses are also present in the ethmoid and maxillary bones.
In comparison, the cranial cavity of humans can be subdivided into three distinct cavities, the anterior, middle, and posterior areas. Since humans are not nearly the keen “smellers ” as most other mammals are, there are no distinct olfactory bulbs, hence no olfactory fossa is present. Further, the bony tentorium of the cat is represented by a fold of the dura mater, the tentorium cerebelli in humans.
THE MANDIBLE
The lower jaw or mandible [Figure 1-22A, Figure 1-22B, and Figure 1-22C] completes the bones of the head. The mandible consists of a pair of dentary bones that articulate cranially at the intermandibular symphysis. Within the first two years of human life the two dentary bones fuse to produce a single bone, the mandible, and unlike the cat, cannot be separated. At the caudal end of the mandible are a pair of condyloid processes, each of which articulates with an elongate groove, the mandibular fossa, in the zygomatic process of the temporal. Notice that these processes are bar-shaped in the cat, a typical design in carnivores allowing them to hold struggling prey but also reducing rotary and lateral grinding movements. The condyloid process of the human is not designed as an elongate bar but is more ovate since this shape is an adaptation of an omnivore, an animal which eats a wide variety of foods. Humans rarely need to subdue struggling prey!
In lateral view, the cranial end consists of a relatively smooth, rounded bar, the body, that contains two or three mental foramina through which pass branches of the mandibular division of
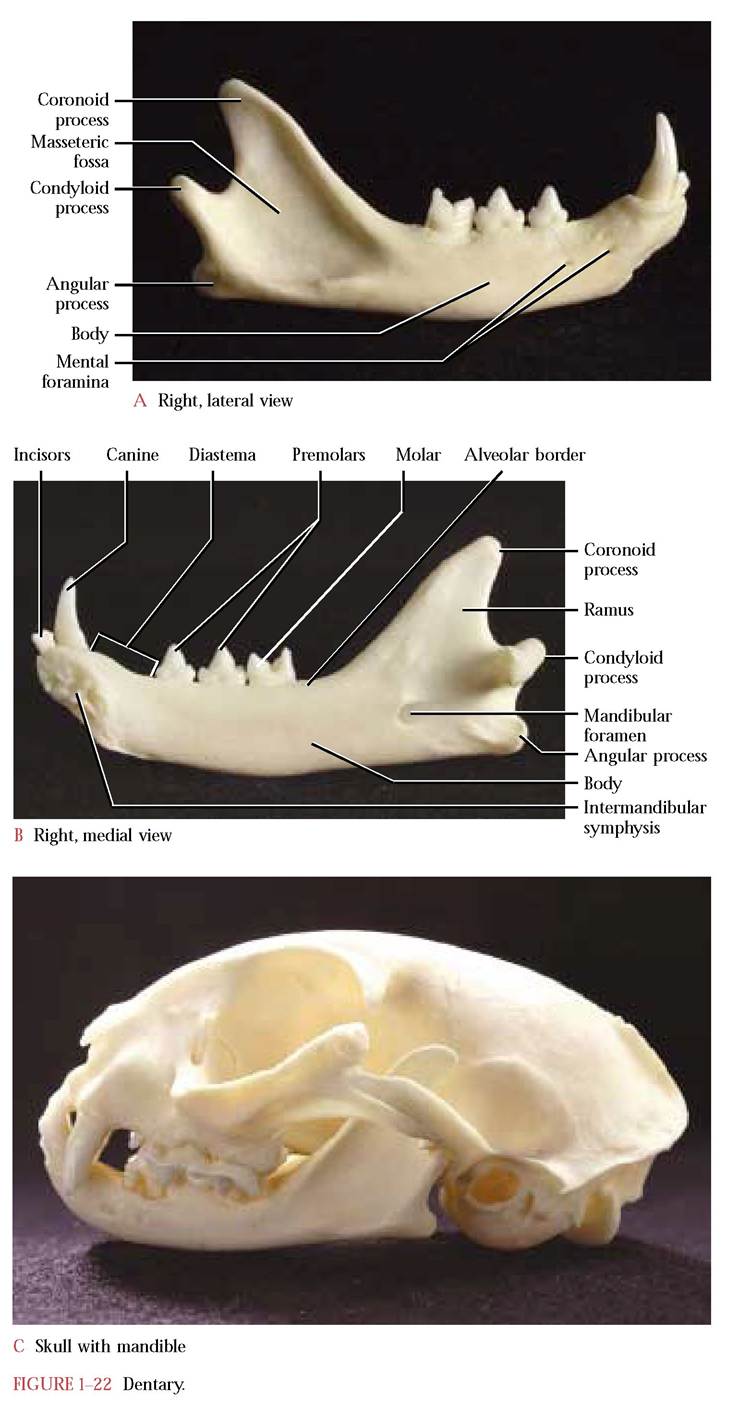
the Trigeminal Nerve (C.N. V). The caudal end of the mandible expands into the ramus. The dorsal part of the ramus, the coronoid process, is the site of insertion of the temporalis muscle, the largest and most powerful of the jaw muscles of carnivores. The well defined triangular-shaped depression, the coronoid or masseteric fossa, accommodates the insertion of part of the masseter muscle, another of the powerful jaw adductors. Notice the small rounded projection, the angular process, at the extreme caudal end of the mandible. Another portion of the masseter and pterygoid muscles insert there.
The medial surface of the mandible is relatively smooth with an exception anteriorly at the symphysis where several rugosities occur to facilitate a strong interlocking joint between the dentaries, and is unevenly sculptured posteriorly where the pterygoid and other jaw muscles insert. The prominent mandibular foramen faces caudally and admits the Mandibular division of the Trigeminal Nerve (C.N. V).
Teeth, anchored in sockets, are found along the dorsal, alveolar border. In each half of the mandible three small
THE VERTEBRAL COLUMN
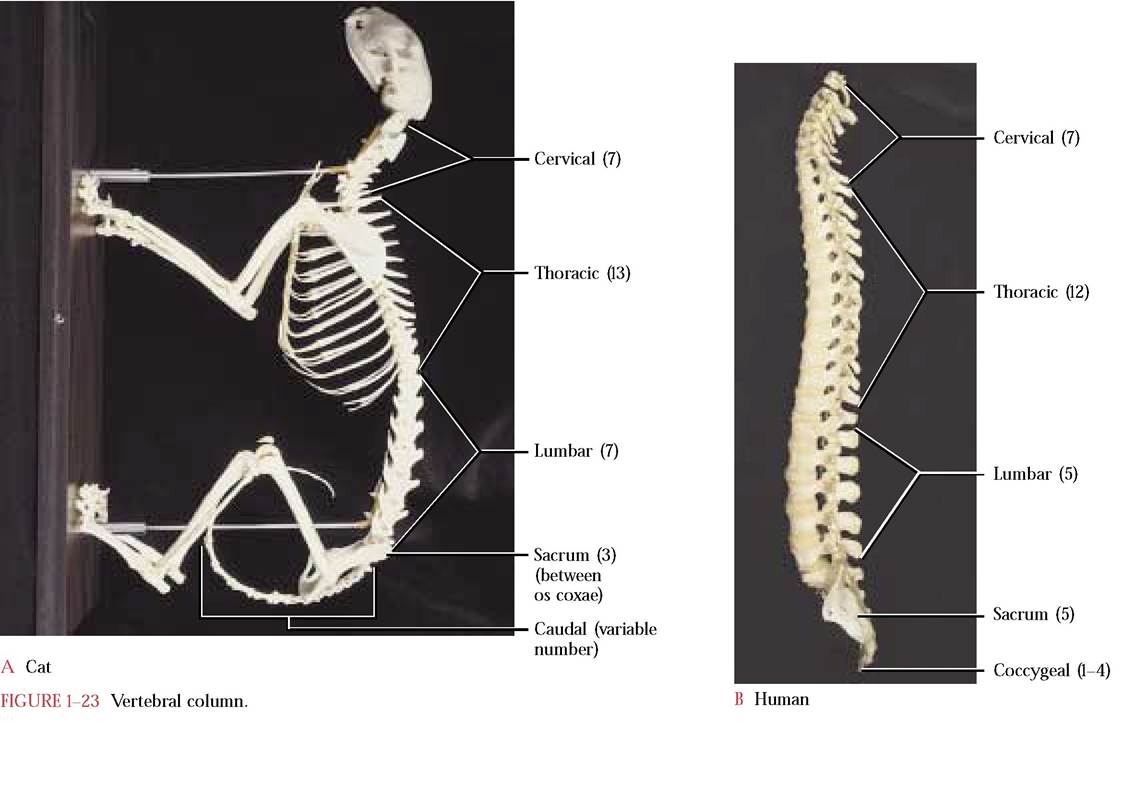
Among terrestrial vertebrates, since body weight is no longer supported by water, mechanical stress caused by the weight of complex organ systems must be transferred to the longitudinal bony beam known as the vertebral column and therefore it has evolved into an important, specialized supporting and locomotory unit. The vertebral column consists of a series of articulated bones called the vertebrae that are distinguishable into five regions: cervical, thoracic, lumbar, sacral, and caudal. The vertebrae provide the main axial support of the body. Support of both appendicular girdles is an essential part of this integrated locomotory and weight bearing unit; the pelvic girdle directly articulates with the sacral region of the vertebral column while the pectoral girdle is associated and held in place by muscles in the thoracic region. The thoracic vertebrae play a secondary and equally important role as the rib-bearing region. Notice in the mounted specimen that the articulated vertebral column has two curvatures, concave in the cervical-thoracic region and
incisors are located anteriorly, followed by a single sharp canine. A wide space, the diastema, separates the canine from the two caudal premolars and a single molar. For further discussion and comparison of the teeth with the human see page 98 of the digestive system. convex in the posterior thoracic-lumbar region that impart bowlike actions to the vertebral column during locomotion. In addition, cushioning fibrocartilaginous intervertebral discs occur between each of the individual vertebrae. If it were not for a concave lumbar curve in the human, the vertebral columns of the cat and human would look very similar [Figure 1-23A and Figure 1-23B].
Cervical Vertebrae
The cervical vertebrae are the most cranial. Almost all mammals—including cats, humans, and giraffes—possess seven cervical vertebrae. Although specializations exist in all regions of the vertebral column, certain features are common to all.
A vertebra is constructed with a solid centrum or body that forms the main ventral support for the spinal cord that rests in the obvious opening, the vertebral canal. The vertebral canal is completed by a dorsal pair of laminae whose dorsocaudal extensions form the spinous process and a pair of pedicles between the laterally projecting transverse processes or diapophyses and the centrum. At the cranial end of each vertebra are two processes; the prezygapophyses, whose articulating facets face dorsally or dorsomedially and articulate with two caudal processes, the postzygapophyses on the posterior end of the vertebra just preceding it. The articular surface of the postzygapophysis faces ventrally or ventromedially. Notice that each vertebra possesses both a pre- and postzygapophysis to facilitate articulation with adjacent vertebrae [Figure 1-24A, Figure 1-24B, and Figure 1-24C].
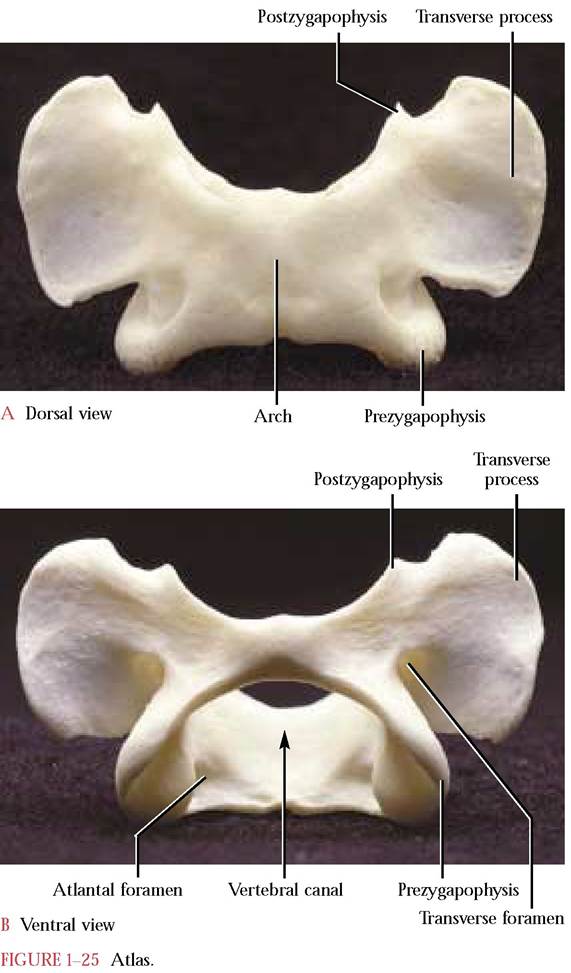
The first and second cervical vertebrae, the atlas and axis, respectively, are very distinctly structured. The ringlike atlas [Figure 1-25A and Figure 1-25B] lacks a centrum and a distinct spine and has broad, winglike transverse processes. The inner articular facets of the expanded pre- zygapophyses accomodate the rounded surfaces of the occipital condyles of the skull. Notice that the postzyga- pophyses do not project as distinctly from the atlas as the prezygapophyses but, nevertheless, bear well defined facets for articulation with the prezygapophyses of the axis.
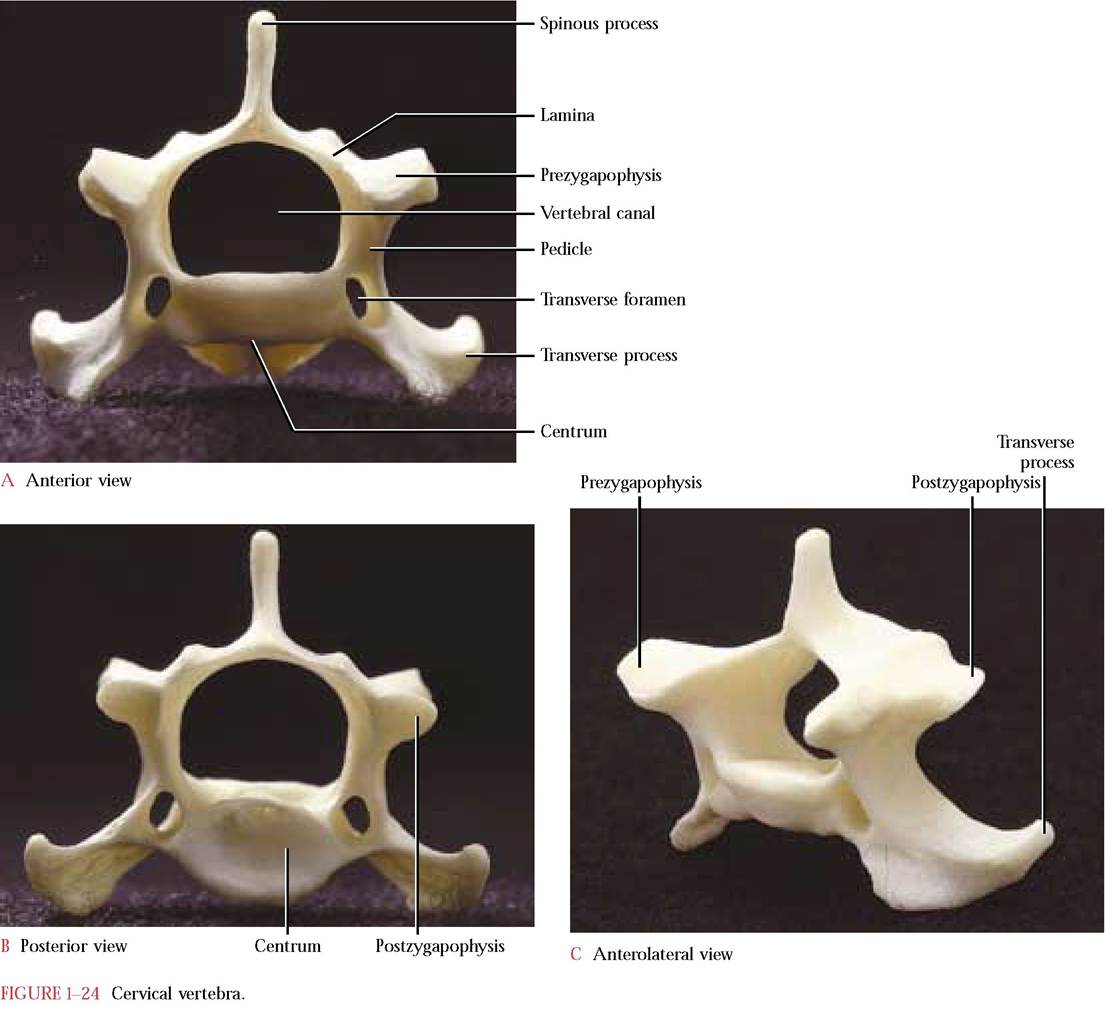
The atlantal foramen, the first intervertebral foramen, through which the first spinal nerve and vertebral vein exit and the vertebral artery enters is located dorsal to the pre- zygapophysis. Typical of the first six cervical vertebrae is the transverse foramen that passes through the transverse process. The vertebral arteries and veins, the blood supply of the brain, pass through these foramina. Look inside the vertebral canal and identify the foramen that allows passage of a small branch of the vertebral artery to form a small artery that extends caudally along the ventral surface of the spinal cord. The human atlas, very similar to that of the cat, possesses only the transverse foramen.
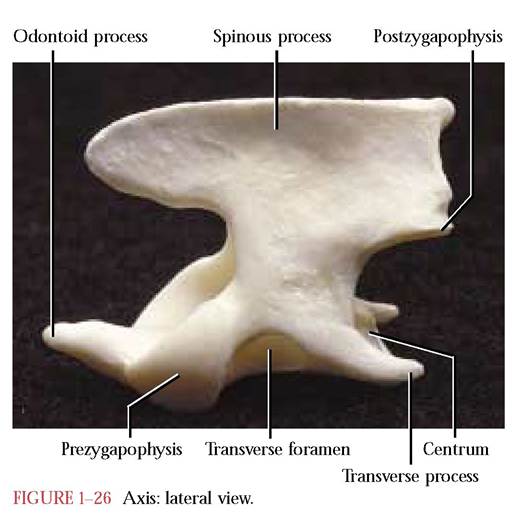
The axis [Figure 1-26], in lateral view, resembles a blacksmith’s anvil with its pronounced spinous process overhanging the arch of the atlas. A second distinctive feature is the cranially projecting odontoid process or dens, laterally flanked by a pair of prezygapophyses with smooth articulating surfaces to articulate with the atlas. The odontoid process represents the centrum of the atlas that has fused with the centrum of the axis and provides an “axle” around which the atlas can pivot to allow rotation of the head on the neck. Take note of the rather typical centrum of the axis in posterior view. Thin caudally projecting transverse processes are present with their characteristic trans-
verse foramina. Find the postzygapophyses with their ventrally directed articulating facets on the dorsal aspect of the arch. Except for a strong reduction of the cranial and caudal portions of the spinous process, the human axis is quite similar to the cat. The human spinous process is bifurcated and strongly grooved posteriorly.
The remaining cervical vertebrae, in many respects, are similar to one another and possess typical vertebral characteristics, with some notable exceptions. The spinous process of the third is very abbreviated, while those of the fourth through the seventh acquire increasing prominence. Note the absence of the transverse foramen in the seventh. The transverse process of the sixth is bifurcated. In humans, cervical vertebrae 2-6 possess short, bifurcated spinous processes, while in the seventh it is single and elongated. All seven cervical vertebrae have transverse foramina.
Thoracic Vertebrae
Among vertebrates, the thoracic vertebrae tend to be the least specialized. Typically, cats have 13 thoracic vertebrae [Figure 1-27]. Identify features that are similar to the cervical vertebrae.
Take note of the articular facet on the ventral surface of the transverse process with which the tuberculum of the rib articulates and also the demifacets occurring on the centrum of two adjacent vertebrae with which the capitulum of the ribs articulates. These facets are unique landmarks found only on thoracic vertebrae.
Neural spines of the first nine or ten thoracic vertebrae are very distinct and elongated, and point caudally whereas the spine of the eleventh or twelfth, the anticlinal vertebra, makes an abrupt change in direction and points cranially. The spines of the rest of the vertebrae posterior to the anticlinal vertebra continue with this orientation. With the anticlinal vertebra there is also a noticeable change in the length of the spinous process. It is in this same area of the vertebral column that accessory processes pointing caudally, appear on the lateral aspects of the pedicles. Note that the eleventh thoracic vertebra has a much reduced transverse process that is absent from the last two vertebrae. The loss of the tuberculum on the last few ribs undoubtedly correlates with the loss of the transverse processes. Among humans there are 12 thoracic vertebrae and all spines project in the same direction and an accessory process, per se, is not present, although it is probably represented by a tubercle on the transverse process.
Lumbar Vertebrae
The seven lumbar vertebrae are characterized as the largest of the vertebrae, increasing in size toward the caudal end [Figure 1-28]. Identify portions of this vertebra that are similar to those of the cervical vertebrae. Perhaps the most obvious landmark on lumbar vertebrae is the cranially projecting pleuropophysis, representing the transverse process with an embryonic rib fused to it. Additionally, accessory processes, toothlike projections, located ventral to the post- zygapophyses, are generally evident on all but the last two, while mammillary processes can be seen on all of the lumbar vertebrae. In the human, there are five lumbar vertebrae that are structurally similar to those of the cat except for the absence of the pleuropophysis.
The Sacrum
This complex consists of three fused vertebrae in the adult,
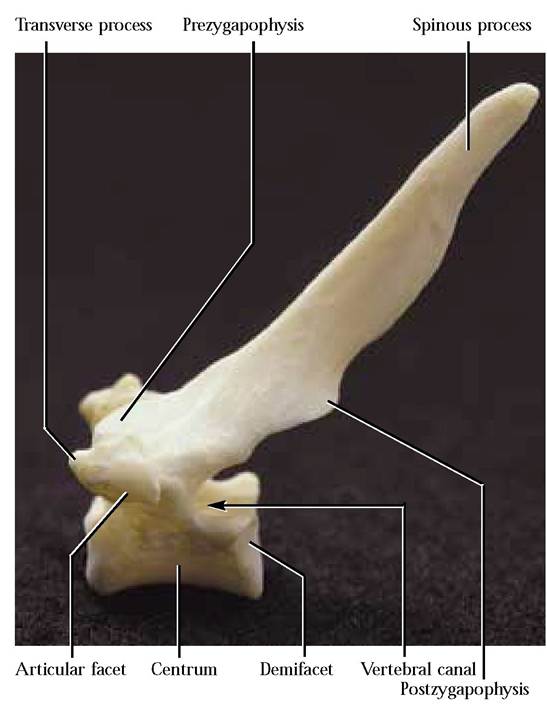
FIGURE 1-27 Thoracic vertebra: lateral view.
but separate in the kitten, that act as a brace for the pelvic
girdle [Figure 1-29A and Figure 1-29B]. Notice that the
three vertebrae decrease in size caudally, while each retains most of the characteristics of the preceding lumbar vertebrae. Note, also, the fusion of the pleuropophyses into a single lateral structure as well as dorsal and ventral foramina to
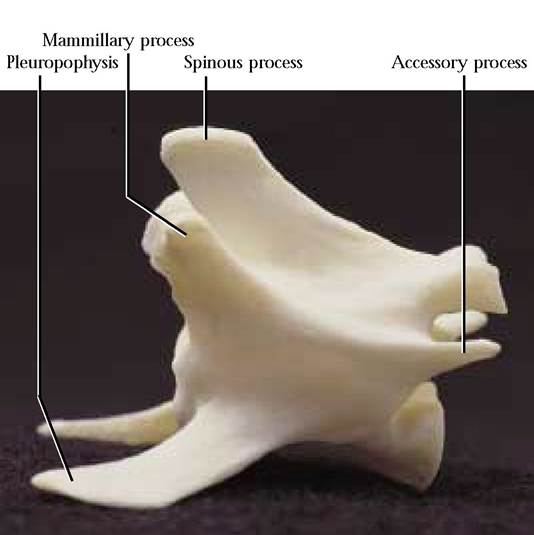
FIGURE 1-28 Lumbar vertebra: lateral view.
Postzygapophysis Dorsal foramen Fused pleurapophyses
Ventral foramina
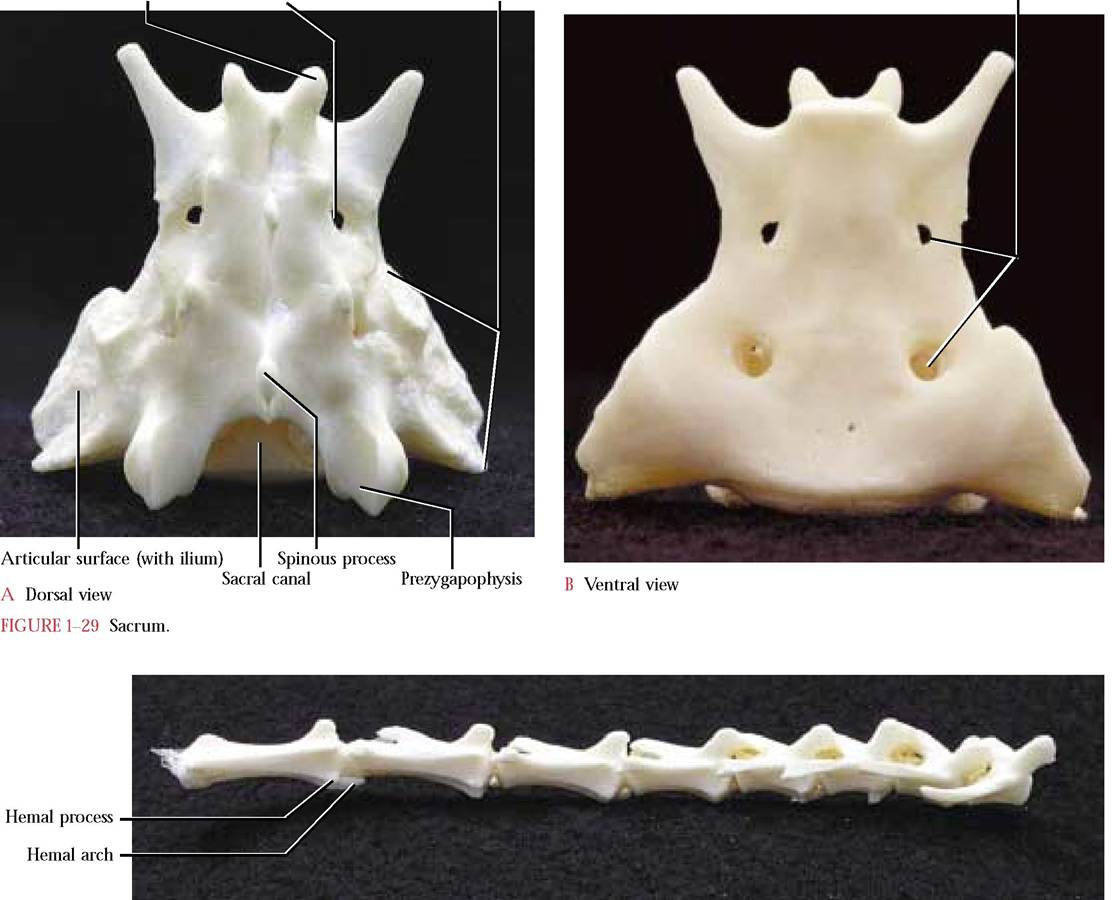
FIGURE 1-30 Caudal vertebrate, anterior toward the right.
accommodate the passage of spinal nerves between each of the adjacent fused vertebrae. The human sacrum is composed of five fused vertebrae and no pleuropophyses.
Caudal Vertebrae
Vertebrae of the tail are the smallest and most variable in number [Figure 1-30]. The more cranial of these exhibit rather typical vertebral characteristics whereas the more caudal vertebrae rapidly lose them and come to resemble simple cylinders representing the centra. Take note of the two small hemal processes on the ventral surface of the centra with which V-shaped hemal arches or chevron bones, housing caudal blood vessels, articulate. Hemal arches identifiable in the intact cat, are often lost during skeletal preparation, but one is present in Figure 1-30. Human caudal vertebrae are much less numerous (3-5), often rudimentary and sometimes fused. These vertebrae are called the coccyx in the human. Usually the only time that we appreciate the fact that we have a tail is when we damage it and are unable to sit comfortably for some time.
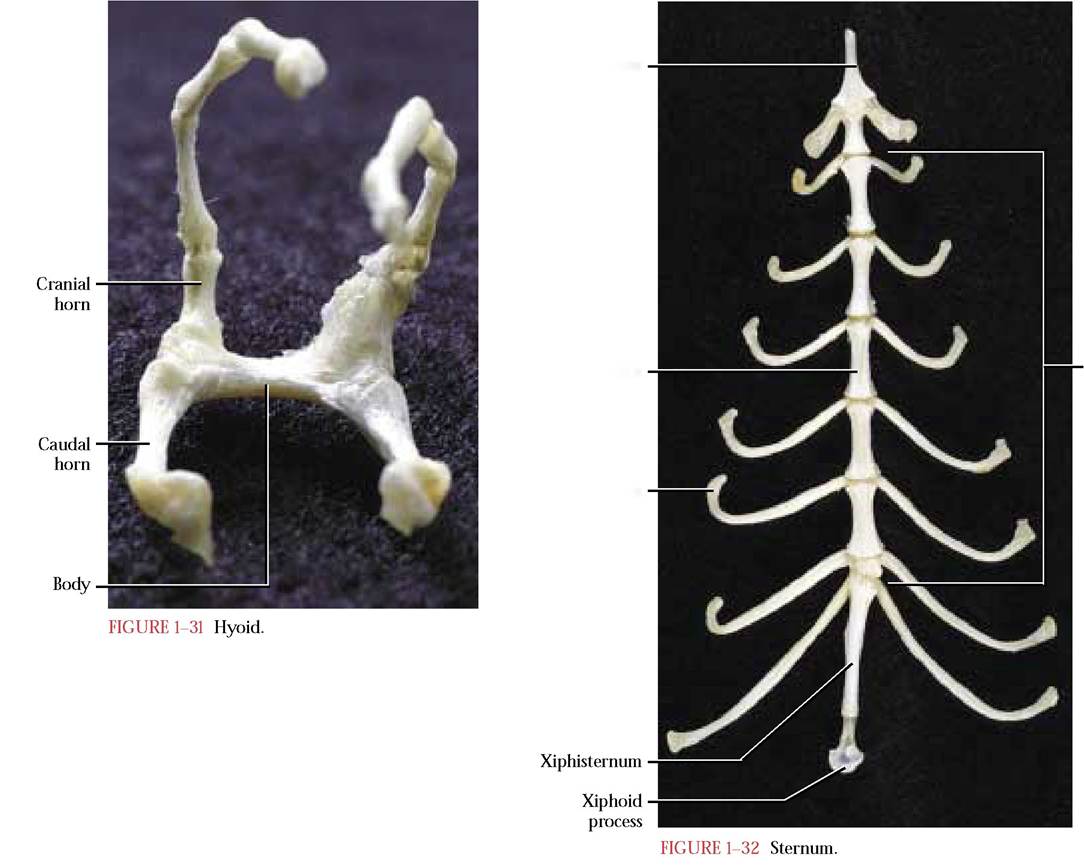
The Hyoid
This is actually a complex of derivatives of several former ancestral gill supports [Figure 1-31]. In mammals, this apparatus, located ventral to the larynx and at the root of the tongue, serves as a site of origin of the tongue and laryngeal muscles. It is H-shaped and consists of a body forming the bar of the H and two cranial or lesser and two caudal or
Manubrium
Sternebra
Costal cartilage
Body
The Sternum
greater horns or cornua, forming the upper and lower uprights of the H. Each cranial horn is connected to the skull by three small bones. In the human, this apparatus is a single fused bone and possesses a body and a pair of greater cornua (caudal horns) and a pair of lesser cornua (cranial horns), similar to the cat.
In the cat, the sternum is slim and elongate and is composed of a series of articulated Sternebrae [Figure 1-32]. It lies ventrally in the midline of the thorax and its caudal end lies just posterior to the diaphragm. It consists of three regions, an anterior slightly keeled manubrium that resembles the tip of a spear, the body consisting of six similar, articulated sternebrae, and a posterior elongated xiphisternum with a distal cartilaginous end known as the xiphoid process.
Costal cartilages associated with the vertebrosternal ribs are generally attached at intervals to the sternum. The first of these is associated with the manubrium at about its midpoint. In some cats, a fusion line indicates the possibility of two individual sternebrae having been united to form the single manubrium. The other eight ribs articulate with the sternum at analogous points between adjacent sternebrae along its length. The eighth and ninth sternal articulations are closely associated with one another. The human sternum is flat and consists of an anterior manubrium, the body representing a fusion of four sternebrae and the terminal xiphoid process whose proximal end may ossify in adulthood. Seven costal cartilages associated with vertebrosternal ribs appear to articulate at positions approximately analogous to those of the cat.
The Ribs
There are thirteen pairs of ribs, the first nine pairs, identified as the vertebrosternal ribs, are considered to be true ribs since their distal ends attach individually to the sternum by means of costal cartilages. There is a great deal of variability in the last four pairs. Generally, the next three pairs or the vertebrochondral ribs, are attached by means of cartilage to each other or to the costal cartilage of the ninth. The final pair has no sternal attachment and therefore is referred to as the vertebral or “floating” ribs.
Although the ribs may be of varying lengths, their morphology is quite similar [Figures 1-33A and 1-33B]. The basic shape is a curved, flattened rod, whose proximal end
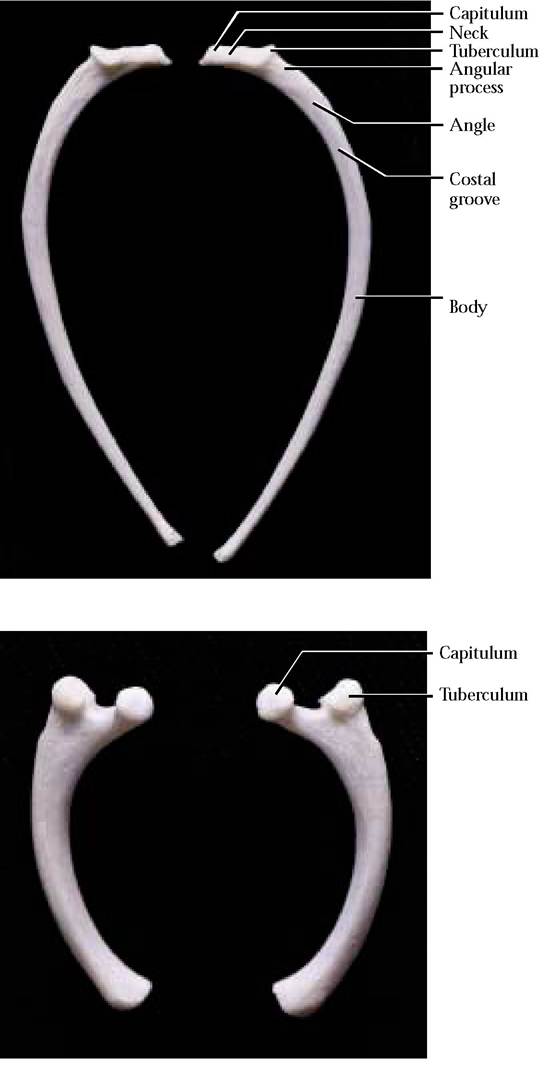
FIGURE 1-33 Left rib, right rib: caudal view.
bears a head or capitulum that articulates with demifacets occurring on the centrum of two adjacent thoracic vertebrae. A second projection, the tuberculum, bearing a smooth facet, articulates with the transverse process of a vertebra. Notice that the tuberculum progressively decreases in prominence antero-posteriorly and is absent from the last two or three pairs of ribs. The slightly constricted area between the capitulum and the tuberculum is known as the neck. The proximal curved portion or angle of the rib blends into the distal part known as the body. A small, pointed projection, the angular process, can be located on the angle of the rib.
Perhaps one of the most confusing aspects of rib morphology is determination of whether it is right or left. One clue is that the articulating surfaces of the capitulum and tuberculum are angled posteriorly, i.e., when examined from the caudal view, the surfaces of the capitulum and tuberculum slant toward you, whereas the opposite is true of a rib looked at from the cranial view. Additionally, a costal groove found along the angle of the rib is oriented caudally. In contrast to the cat, humans possess twelve pairs of ribs, seven of which are vertebrosternal, three vertebrochondral and two vertebral.
Appendicular division
Pectoral Girdle and Appendage
The Clavicle
The clavicle in cursorial (running) mammals is drastically reduced and often embedded in the muscle of the shoulder. In the cat, it is a slender, rod-like, curved bone with the sternal end slightly enlarged [Figure 1—34]. The human clavicle is much more robustly constructed with a slight sigmoid curve and its articulation reflects a more primitive mammalian condition. It articulates medially with the manubrium of the sternum and laterally with the acromion process of the scapula.
The Scapula
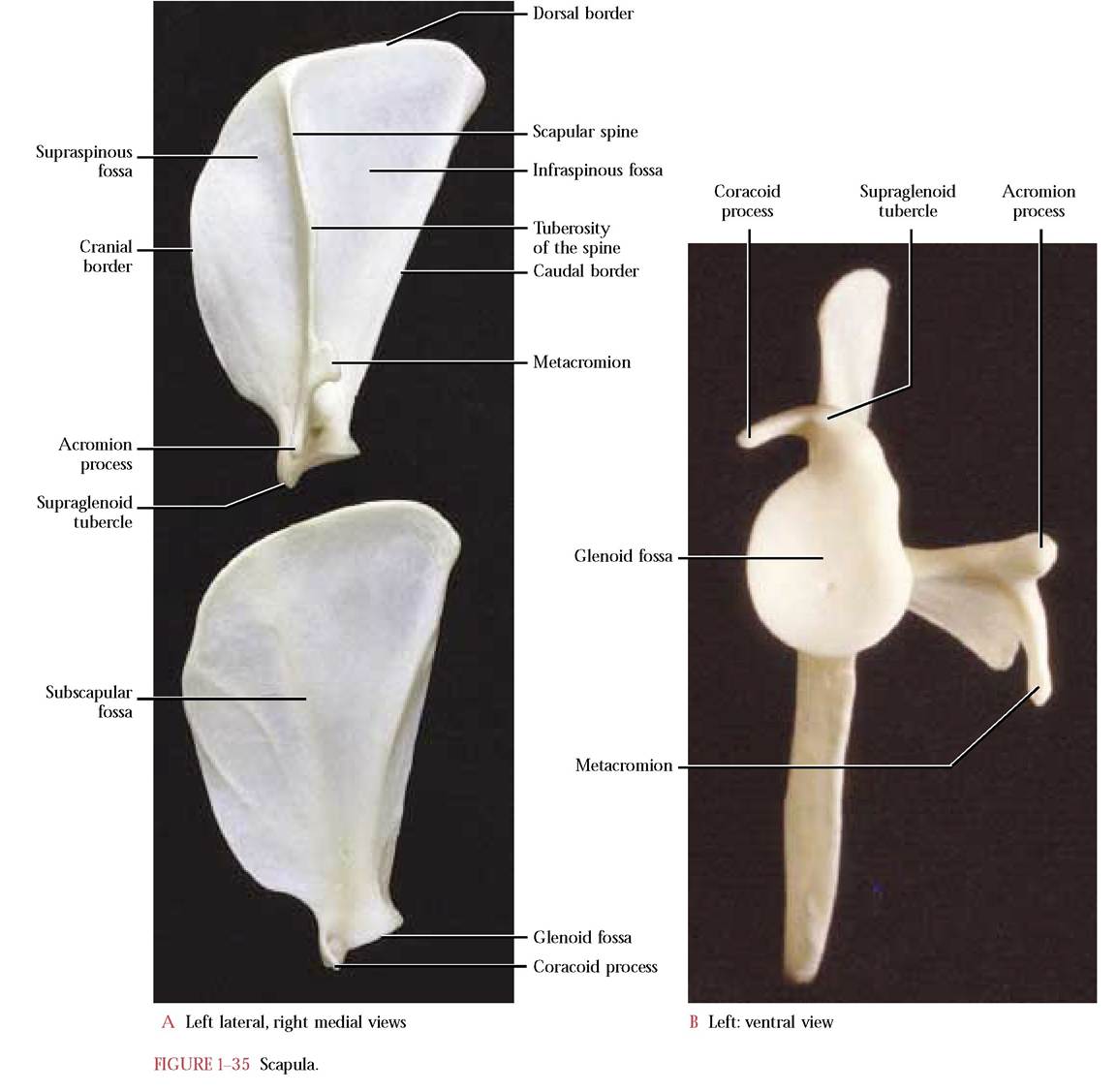
The scapula is a flat triangular bone that articulates with the humerus [Figure 1-35A and Figure 1-35B]. The curved dorsal or vertebral border continues anteriorly as the cranial border and posteriorly as the caudal or axillary border. At the vertex of the triangle is a concave surface known as the glenoid fossa with which the head of the humerus articulates. A small beak-like projection, the coracoid process, extends medially from the anterior border of the glenoid fossa. Lateral to it is the supraglenoid tubercle. A prominent lateral ridge, the scapular spine, begins at about the midpoint of the vertebral border and extends just dorsally to the glenoid fossa and terminates in a free, pointed acromion process. The upper curved, thicker portion of the free edge is identified as the tuberosity of the spine while the caudally projecting, sharp-angled metacromion occurs just dorsal to the acromion. The spine divides the lateral surface into an anterior supraspinous fossa and a posterior infraspinous fossa.
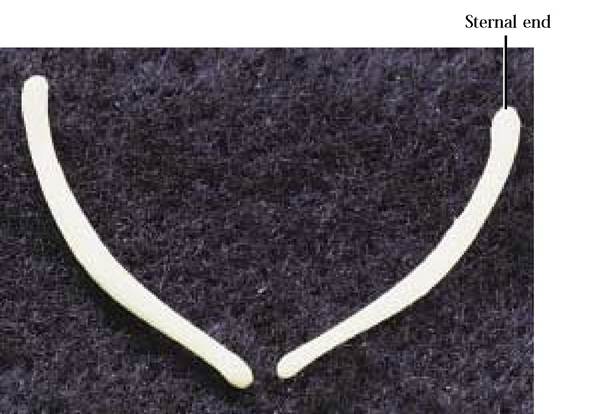
FIGURE 1-34 Right and left clavicle: cranial view.
On the medial surface, the subscapular fossa, is somewhat flat with a narrow elevation marking the medial line of the spine overlying it on the lateral surface. Two oblique ridges are obvious landmarks indicating points of muscle attachment on the anterior surface of the fossa, while a single prominent ridge occurs along the posterior margin of the fossa again indicating the point of muscle attachment. A somewhat concave surface is present between this ridge and the posterior margin for muscle attachment. A variable number of nutrient foramina are present on both surfaces. Other than the disproportionate surface area of the infraspinous portion of the scapula in the human, the cat and human scapulae are quite similar.
The Humerus
The proximal or upper long bone of the anterior limb is known as the humerus [Figure 1—36]. It articulates proximally with the scapula and distally with the radius and ulna. It consists of a slightly curved diaphysis and two prominent epiphyses. The proximal epiphysis consists of a smooth, rounded medial head that articulates with the glenoid cavity of the scapula. Medial to the head is the smaller lesser tuberosity and lateral to the head is the larger greater tuberosity for muscle attachment. The bicipital groove, that accommodates the tendon of the biceps brachii muscle, separates these two tuberosities. At the distal end are a pair of prominent condyles, a larger medial trochlea and a smaller lateral capitulum, with which the ulna and radius articulate, respectively.
Proximal to the trochlea is the coronoid fossa and proximal to the capitulum is the radial fossa. Medial and proximal to the trochlea is the prominent medial epicondyle and lateral and proximal to the capitulum is the less obvious lateral epicondyle. The very conspicuous ovoid slit occurring proximal to the medial epicondyle is the supracondyloid foramen. Through the foramen passes the median nerve and the brachial blood vessels. On the posterior side proximal to the trochlea and capitulum is the deep, prominent olecranon fossa with which the olecranon of the ulna articulates.
Posteriorly, a sharp supracondyloid ridge extends from
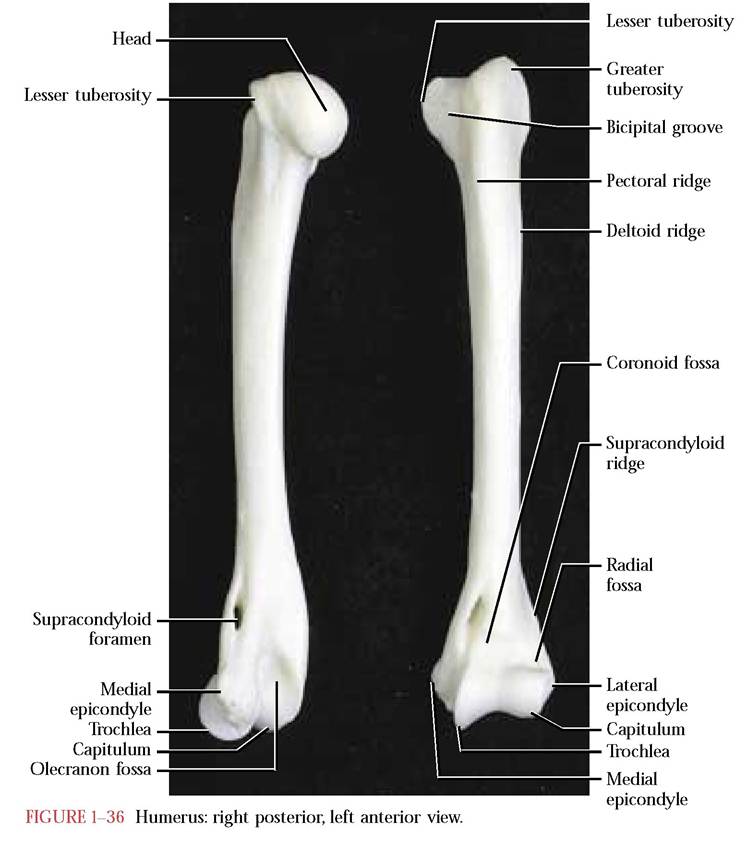
the lateral epicondyle to about the midpoint of the diaphysis. A prominent rugosity, the medial pectoral ridge, and the sharp, crestlike, lateral deltoid ridge converge anteriorly at about midshaft. These surface irregularities mark the positions of muscle attachments. Numerous nutrient foramina occur in both the diaphysis and the epiphyses of the humerus. Other than the presence of the anatomical neck and the absence of the supracondyloid foramen, the human humerus is similar to the cat.
The Radius
The natural position of the radius [Figure 1-37] in the forelimb of the cat extends from the lateral proximal humeral end to the medial distal carpal end, therefore this long bone crosses over the ulna. As is typical of a long bone, it consists of a central diaphysis and proximal and distal epiphyses. The proximal epiphysis or head is slightly concave to accomodate its articulation with the capitulum of the humerus. Circumscribing the head is the narrow, smooth articular circumference for articulation with a notch in the ulna. Below the head is the neck. Just distal to the neck on the postero-lateral surface of the diaphysis is the bicipital tuberosity for the insertion of the tendon of the biceps brachii muscle. Distal to the
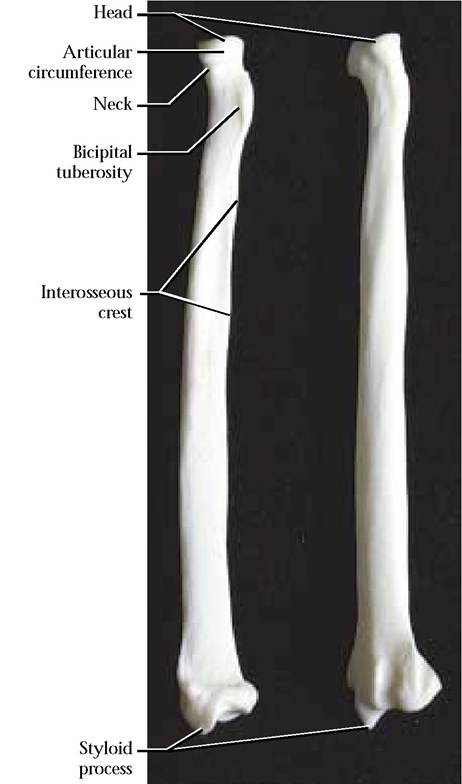
FIGURE 1-37 Radius: right posterior, left anterior view.
bicipital tuberosity, is an obvious roughened surface, the interosseous crest. The distal epiphysis is characterized by the presence of the medial styloid process. Just proximal to the styloid process is a concavity with which the radius articulates with the scapholunate of the carpals. On the lateral surface is a small concave facet with which the ulna articulates. On the anterior surface exist several grooves separated by prominent ridges to allow passage of tendons. The anatomy of the human radius is very similar.
The Ulna
The second bone of the forelimb is the ulna, another long bone consisting of a diaphysis and a pair of epiphyses [Figure 1—38]. The proximal end is better known as the olecranon which articulates with the olecranon fossa of the humerus. Furthermore, this process functions as the insertion site of the tendon of the powerful forearm extensor, the triceps brachii. On the anterior surface, just distal to the olecranon is an unusually shaped, articulating surface, the deeply excavated semilunar notch. The trochlea of the humerus articulates with the notch. An anterior projection, the coronoid process, forms the medial margin of a lateral concave articulating surface, the radial notch, for the articulation of the head of the radius.
The distal end of the ulna is distinguished by the peg-like lateral styloid process. The medial surface articulates with the cuneiform and pisiform of the carpals. On the medial surface of the ulna, just proximal to the styloid process, observe the small articulating surface for the articulation of the radius. Along the antero-lateral aspect of the ulnar diaphysis, notice the usually prominent interosseous crest that approximates the crest on the corresponding area of the radius. A tough sheet of connective tissue extends from
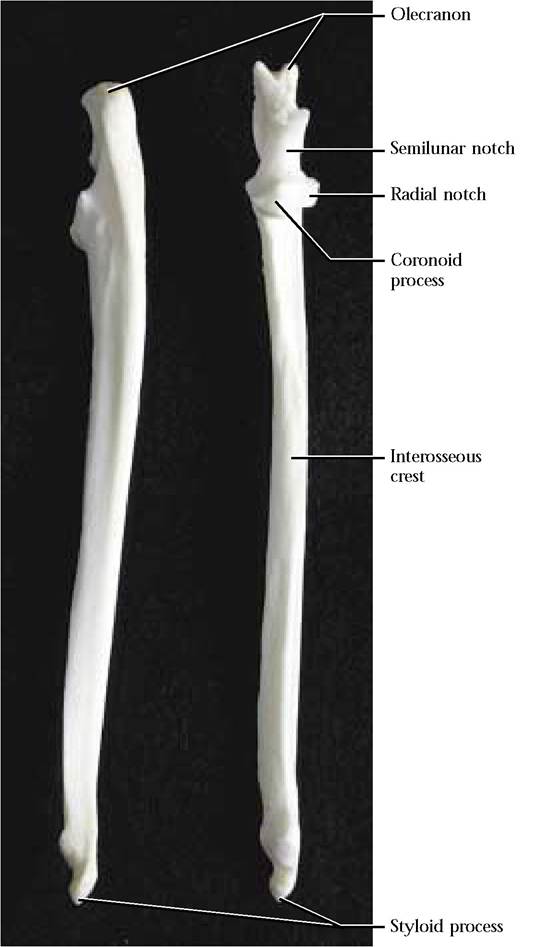
FIGURE 1-38 Ulna: right posterior left anterior view.
the crest of the radius to the crest of the ulna, stabilizing the lower foreleg by preventing major torsion or twisting movement between the radius and ulna. The human ulna is very similar in structure to the cat.
The Carpals and the Manus
A series of seven, small, irregularly shaped bones, the carpals, organized into two rows, a proximal set of three and a distal set of four, separate the forefoot or manus from the foreleg and are known as the wrist bones [Figure 1—39].
The most medial and largest of the proximal row is the fused scaphoid and lunate called the scapholunate. In the human, these two bones occur as separate carpals. In the middle of this row is the cuneiform. This carpal is known as the triquetral in the human. The most lateral of the proximal set is called the pisiform. The human pisiform is similar. Although from the anterior view the pisiform appears as a small bone, a considerable portion projects posteriorly, therefore, it is substantially larger than it first appears. This
In the distal row, from medial to lateral, are the trapezium, trapezoid, capitate, and hamate. These are similar in the human. Distally, these bones articulate with the metacarpals of the forefoot or manus.
The manus consists of five metacarpals with which the proximal phalanges of the five digits articulate [Figure 1-39]. There are two phalanges in the thumb and three in each of the other four toes. Sesamoid bones occur at the junctions of the metacarpals and the phalanges. The cat, typical of most feline carnivores, has well-developed retractile claws, an obvious adaptation among these predators to aid in capturing and holding frisky prey. Human digits are covered by flat nails.
Pelvic Girdle and Appendage
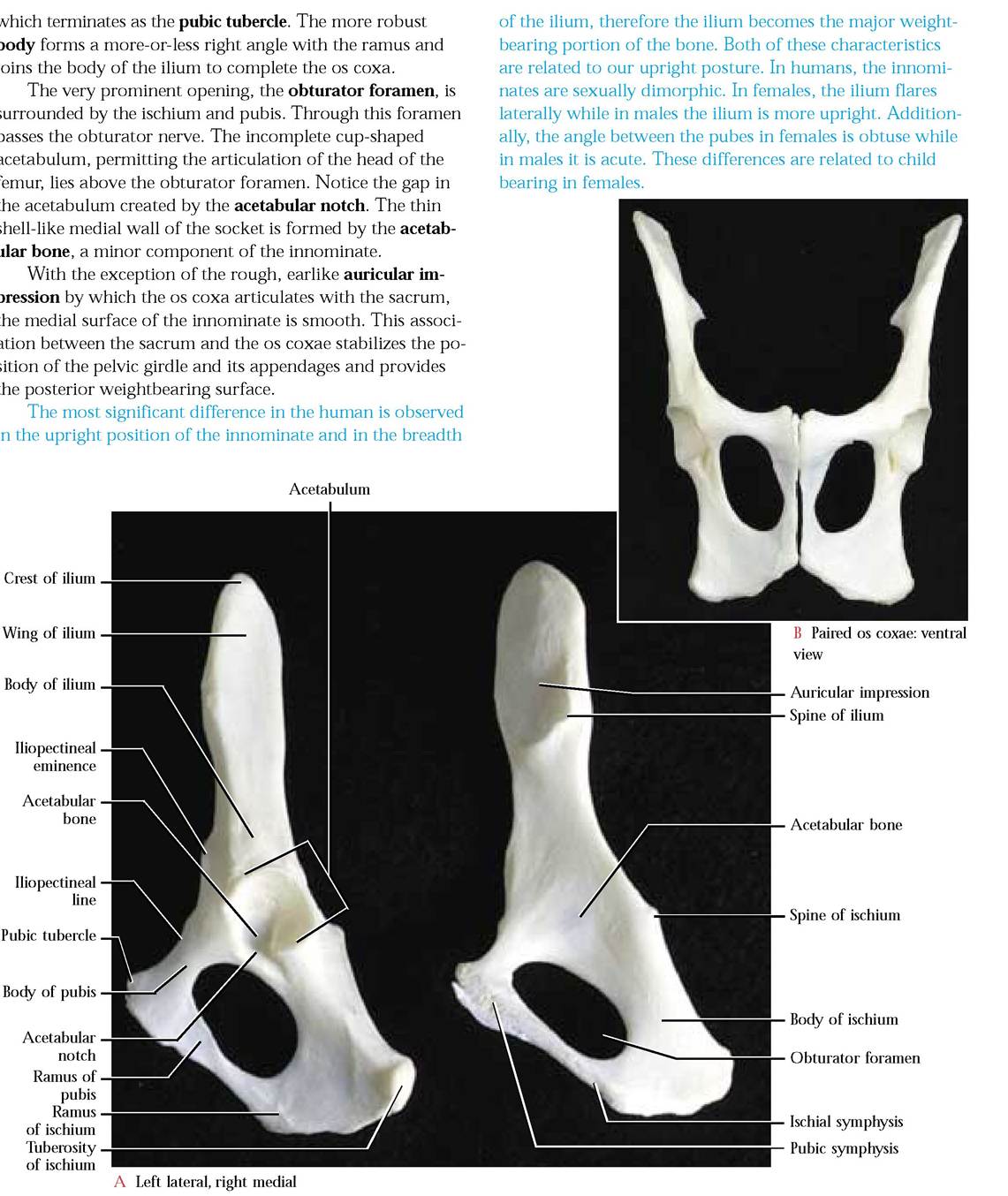
The Os Coxa or Innominate
This irregularly shaped, elongated, complex consists of three individual bones, the ilium, the ischium, and the pubis [Figure 1-40A and Figure 1-40B]. In the juvenile cat, these
row articulates proximally with the radius and ulna and distally with the second row of carpals. bones can be distinguished because they are separated by sutures. In the adult cat they become fused and the sutures become less obvious. The pelvic girdle of the cat is comprised
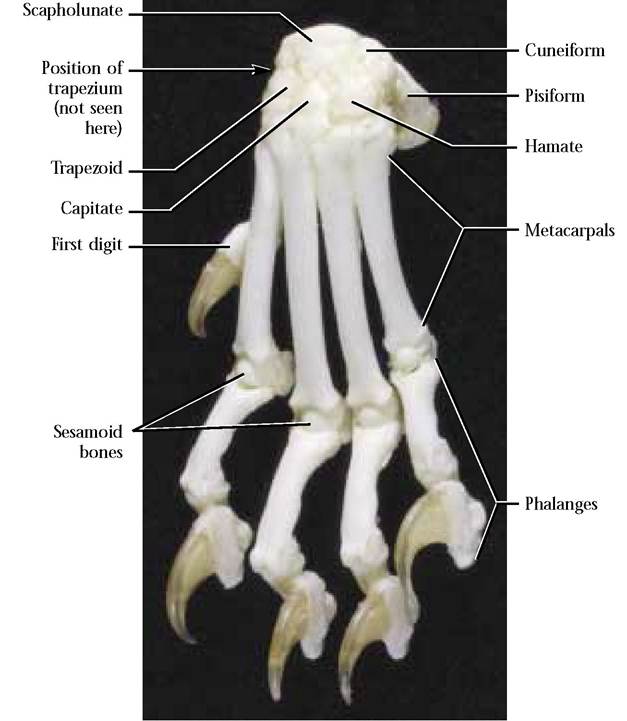
FIGURE 1-39 Left manus and carpals.
of the paired innominate bones that articulate ventrally along the medial surfaces of the pubic and ischial portions forming the pubic and ischial symphysis, respectively [Figure 1-40B]. The ilial portion of the innominates is rather firmly articulated medially with the sacrum, a specialized region of the vertebral column.
The cranially projecting ilium is wing-shaped with a concave lateral surface and is called the wing of the ilium. The dorsal portion of the wing is roughly thickened and is called the crest. The thickest portion of the ilium proximal to the wing is the body and its base contributes to the formation of the deeply concave acetabulum. The antero-ventral margin of the ilium is known as the ileopectineal line. This pelvic landmark continues onto the pubic surface to the symphysis. At about the level of the acetabulum lies the ileopectineal eminence along the ileopectineal line. Along the posterior border below the crest is the spine of the ilium.
The posterior part of the ischium consists of the heavy, broad body and the more lightly constructed, ramus. The roughened, thickened posterior end of the body is known as the tuberosity of the ischium. Very near the cranial end of the ischium and along the posterior border is the fairly prominent spine of the ischium.
The third bone that contributes to the formation of the innominate is the pubis. That portion of the pubis that is continuous with the ramus of the ischium is logically called the ramus of the pubis
The Femur
The proximal bone of the posterior limb is the femur [Figure 1-41]. It is a typical long bone consisting of a central shaft or diaphysis and a proximal and distal epiphysis. A prominent ball-like projection, the head, with an irregular depression, the fovea capitis, is located on the medial aspect of the proximal epiphysis. The head articulates with the acetabulum of the innominate and the fovea capitis is the site of attachment of a ligament that helps anchor the femur to the ventral portion of the acetabulum. The constricted neck connects the head to the epiphysis. Equally as prominent on the lateral aspect of the femur is the greater trochanter where hip muscles attach. Medial to the greater trochanter is a conspicuous depression, the trochanteric fossa. Extending
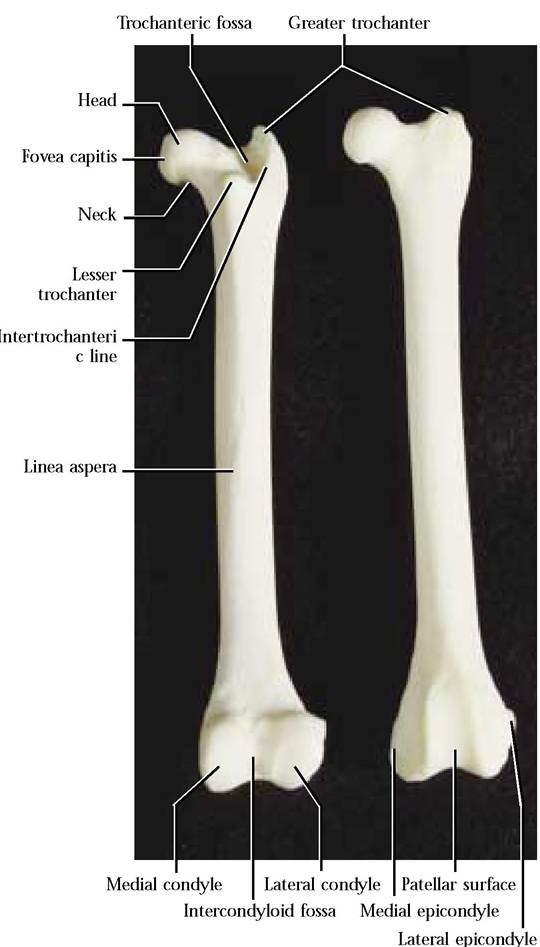
FIGURE 1-41 Femur: right posterior, left anterior.
from the greater trochanter and running along its rim and terminating in a small triangular projection, the lesser trochanter, is the intertrochanteric line. Several ridgelike lines extend around the neck and lesser trochanter. Notice the prominent ridge extending from the greater trochanter and along the postero-lateral aspect of the femur, crossing from lateral to medial at midshaft. This ridge is joined by a much less conspicuous line extending from the lesser trochanter to form the linea aspera. These ridges provide sites of muscle attachment.
The distal end of the femur is distinguished by two prominent projections, the medial and lateral condyles whose smooth, rounded surfaces articulate with the proximal end of the tibia. A deep posterior notch, the intercondyloid fossa, separates the condyles. Take note of the two irregularly shaped prominences, the medial and lateral epicondyles, located above the condyles and providing sites for muscle attachment. The anterior tongue-shaped, smooth surface, the patellar surface, is joined by the patella (“kneecap”) to form a smooth surface over which major extensor muscle tendons ride. Several very conspicuous openings, nutrient foramina, for the passage of blood vessels and nerves are found in the diaphysis and both epiphyses. Generally, the human femur is very similar.
The Patella
The kneecap or patella [Figure 1-42] is a major member of that group of bones known as sesamoid bones that are associated with muscle tendons, in this case the tendon of the quadriceps femoris muscle, the major extensor of the leg. Its shape resembles a smooth pumpkin seed and its posterior surface articulates with the condylar articular surfaces of the femur and therefore appears as a negative mirror image of the patellar surface of the femur. The human patella is very similar to the cat patella.
The Tibia
The tibia [Figure 1-43] is the longer and more robust of the two lower leg bones and is the primary weightbearing bone of the hindlimb. It consists of a triangular diaphysis and two irregular epiphyses. The proximal epiphysis is roughly triangular, when viewed from above. The base of the triangle includes a medial and lateral condyle whose surfaces exhibit articular facets that accomodate the articular surfaces of the femoral condyles. The popliteal notch on the posterior surface delineates the two condyles. On either side of the condyles are a medial and lateral tuberosity. A double pointed projection, the spine separates the articular facets. A triangle, whose base is formed by the medial and lateral tuberosities, has its vertex marked by the tibial tuberosity. Continuous with the tuberosity is the tibial crest extending along the anterior surface of the femur. Beneath the lateral condyle is the articular facet for the head of the fibula.
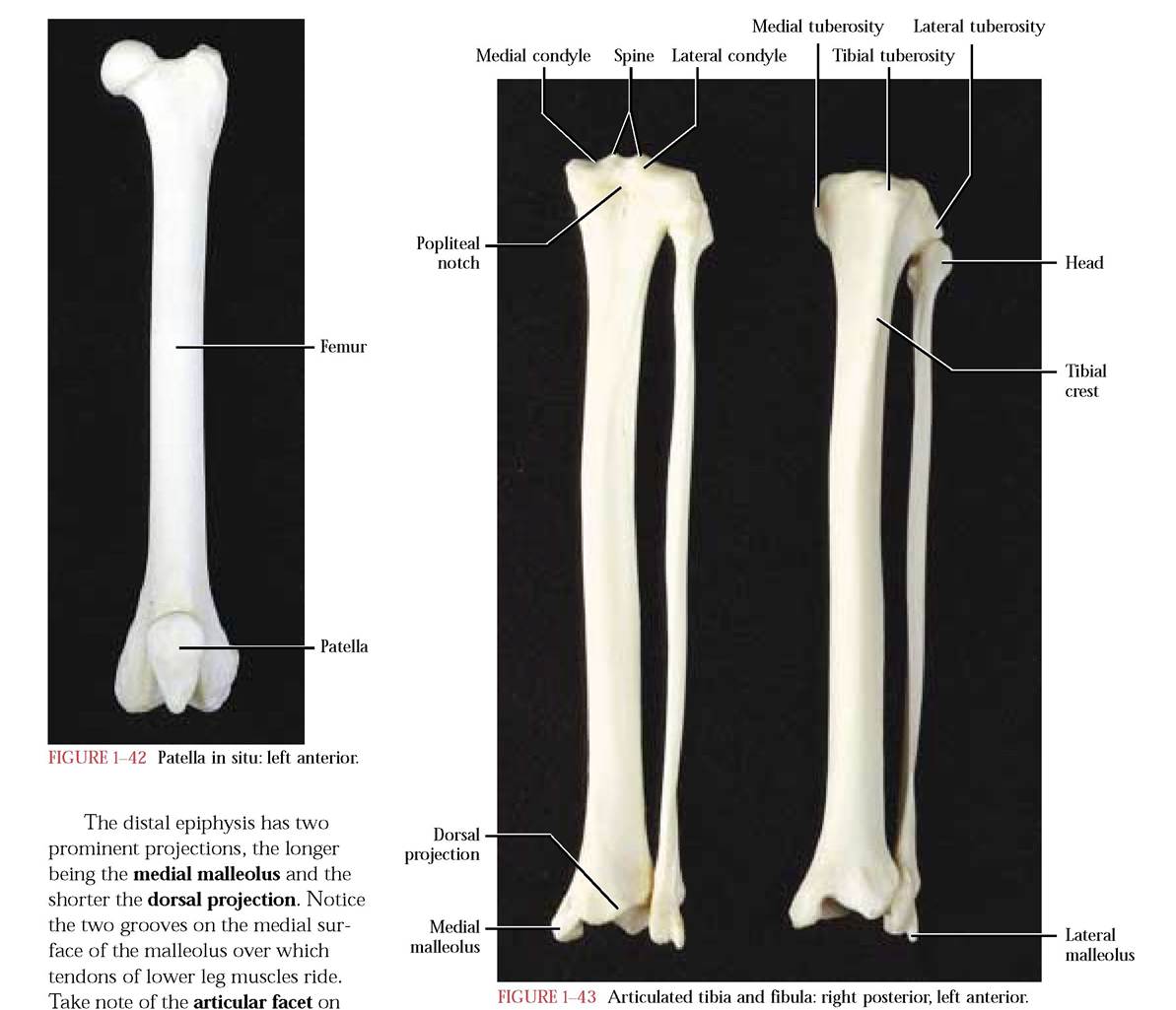
the postero-lateral surface of the dor
sal projection for articulation with the fibula. The lower surface of this epiphysis consists of two concave articular facets with which the talus (astragalus) of the ankle articulates.
Several ridges identifiable as muscle attachment regions can be observed on the posterior surface of the diaphysis. Morphologically, the human tibia is very similar to that of the cat.
The Fibula
The slender long bone of the hind limb, lateral to the tibia is the fibula [Figure 1—43]. Similar to other long bones it consists of a diaphysis and two epiphyses. The proximal epiphysis is comprised of an irregularly shaped head whose medial surface bears a smooth facet for articulation with the lateral surface of the tibia. The medial surface of the diaphysis is flattened, while the lateral surface is gently curved. The distal epiphysis is distinguished by the lateral malleolus. Facets for articulation with the tibia and talus can be found on its medial surface. Grooves to accomodate lower leg muscle tendons are visible laterally and ventrally. The human fibula is very similar.
The Tarsals and the Pes
The number of tarsal bones (7) is identical to the number of carpals, however, in contrast to the carpals, they vary in
size [Figure 1-44]. This is particularly true of the medially located talus whose size and shape probably relate to the fact that it is the bone with which the tibia and fibula articulate and therefore represents the weightbearing bone of the ankle set. The calcaneus, laterally located, is the longest bone of the ankle and forms the heel. The groove on the exposed surface of the calcaneus accommodates the tendon of Achilles. Distal to the talus lies the navicular. A row of four tarsal bones, identified from medial to lateral, consist of the medial cuneiform, intermediate cuneiform, lateral cuneiform, and cuboid.
The pes consists of five metatarsals that articulate with the distal row of four tarsal bones. With the exception of the first, which has been greatly reduced, the other four are very elongated and each articulates with a series of three phalanges, and similar to the manus terminate in a well developed retractile claw [Figure 1-44]. Note the Sesamoid bones at the junction of the metatarsals and phalanges.
There are two major differences in the human. One of these is the presence of the first digit or “big toe” and the second is the way in which the human walks on its foot, since the entire foot from heel to toe is placed on the substrate. In walking on its toes, the cat has increased its leg length, reduced the amount of friction between the foot and substratum, therefore enabling it to run swiftly.
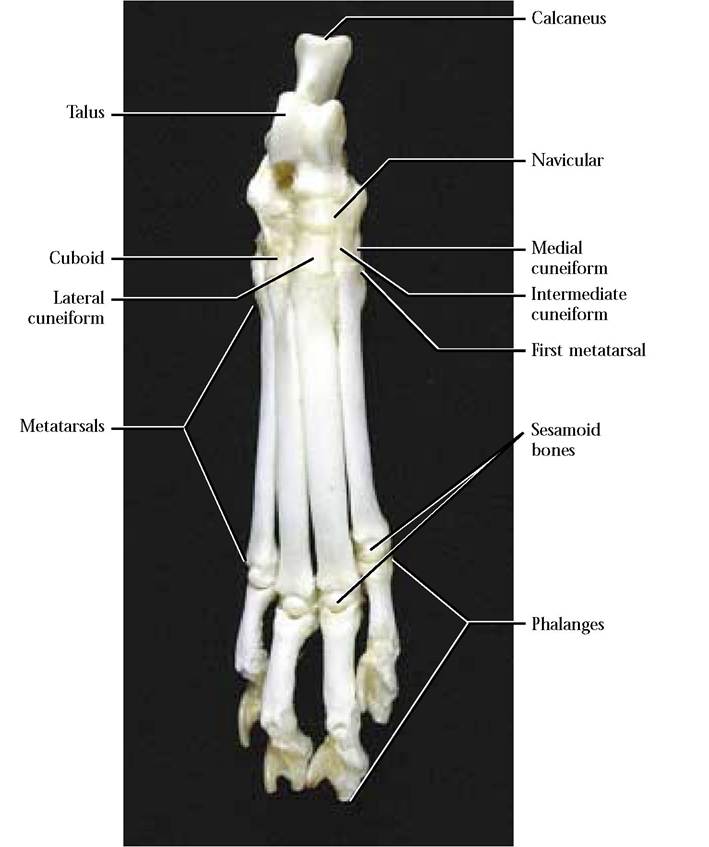
FIGURE 1-44 Right pes and tarsals.