Somatic Motor Pathways
Somatic motor activity is regulated hierarchically within the central nervous system by separate groups of neurons termed the lower motor neurons and upper motor neurons.
Lower motor neurons are neurons that have cell bodies located within the central nervous system and axons that leave the central nervous system to synapse upon muscle fibers in the periphery— these are the neurons that have been named motor neurons up to this point.
The axons of lower motor neurons form the motor component of peripheral nerves. The cell bodies of those lower motor neurons, which innervate the skeletal muscle of the neck, trunk, and limbs, are located within the ventral portion of the gray matter of the spinal cord (Figs. 8.12/4 and 8.13/6) and their axons travel in the peripheral nerves of the neck, trunk, and limbs (Fig. 8.48). The cell bodies of those lower motoneurons that innervate the skeletal muscle of the head are located within somatic motor nuclei in the brainstem, and their axons travel in the cranial nerves that contain somatic efferent components. The number of lower motor neurons innervating a particular muscle varies with the precision of performance required of that muscle (p. 23). Lower motor neurons are the efferent component of local reflex responses that involve the muscles in question (Fig. 8.48) but are also importantly controlled by the activity of upper motor neurons.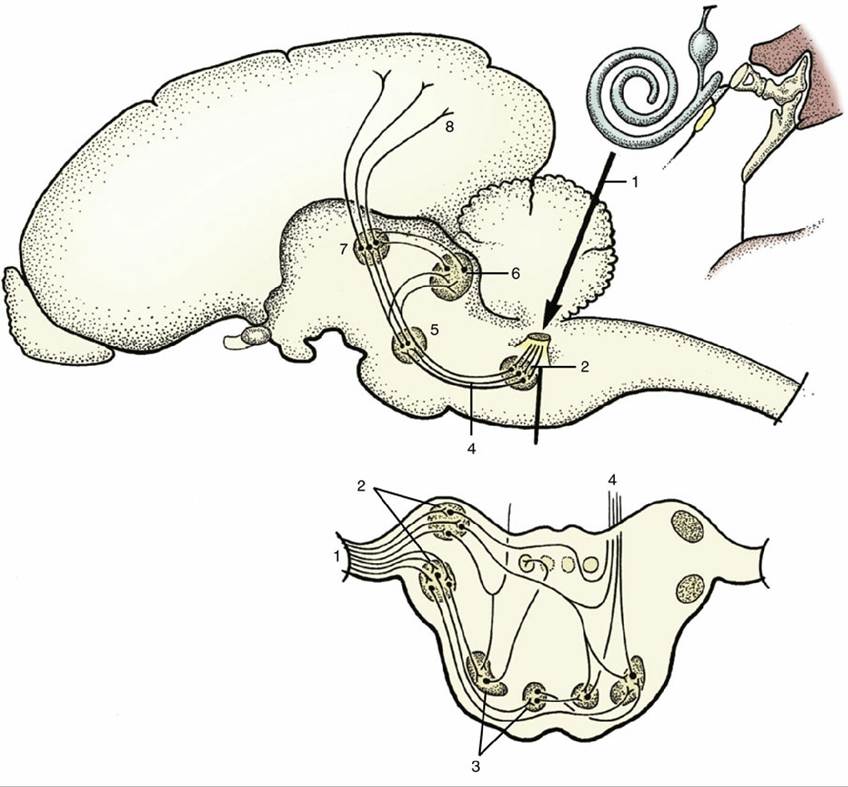
FIG. 8.47 A simplified scheme of the auditory pathways. 1, Cochlear fibers in the vestibulocochlear nerve; 2, cochlear nuclei (dorsal and ventral); 3, nuclei in trapezoid body; 4, lateral lemniscus; 5, nucleus in lateral lemniscus; 6, caudal colliculus; 7, medial geniculate nucleus; 8, projection fibers for conscious perception.
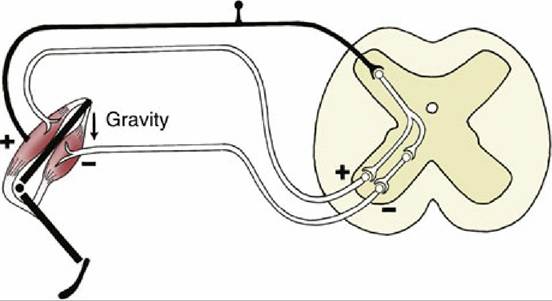
FIG.
8.48 A myotactic reflex arc. Gravity (arrow) stretches the extensor muscle, stimulating its contraction via the reflex arc. To allow shortening of the extensor muscle, the flexor muscle is inhibited at the same time by a collateral fiber and an inhibiting interneuron.The upper motor neurons are involved in more complicated reflexes and also initiate voluntary movements. The cell bodies of upper motor neurons are located in the brain, within the motor area
of the neopallium but also in other regions of the brain, including the reticular formation and red nucleus. The axons of these neurons travel through the brainstem and spinal cord to synapse on lower motor neurons. The amount of cortical area allocated to upper motor neurons controlling the lower motor neurons and, ultimately, the muscles of different parts of the body vary in extent with the importance and complexity of the movements of these parts in the habitual activities of the species. Thus the region of the cortex controlling the hand in humans is relatively much larger than that allocated to the whole limb in ungulates. Importantly, the axons of upper motor neurons do not leave the central nervous system and thus do not project directly on muscle fibers but exert their control by excitation or inhibition of lower motor neurons.
The axonal connections between the upper and lower motor neurons follow pathways that vary considerably among species in their relative development and details of organization. The primary distinction, derived from human neuroanatomy, is made between so-called pyramidal and extrapyramidal systems, although the two are coordinated and work in close collaboration. The pyramidal system is mostly concerned with the exercise of finely adjusted movements, whereas the extrapyramidal system is employed in the control of coarser movements, particularly in stereotyped locomotor patterns. It follows that the pyramidal system is better developed in primates than in domestic species, a distinction that explains the different consequences of lesions to the pyramidal pathway.
Severe damage to the pyramidal pathway produces a complete and permanent paralysis of the contralateral voluntary musculature in primates, but the effects in domestic species are mainly confined to disturbance of contralateral postural reactions from which partial recovery occurs after a few days. Both pyramidal and extrapyramidal systems are provided with elaborate feedback mechanisms that allow for the continuous monitoring and adjustment of motor activity.The Pyramidal System
The pyramidal system takes origin from upper motor neurons within various regions of the neopallium, particularly the primary motor area. The axons of these neurons converge as they exit the telencephalon and form an important portion of the internal capsule; in their passage they preserve the orderly point-to-point arrangement of the cortical motor representation. They then continue over the lateral aspect of the thalamus to enter the crus cerebri on the ventral surface of the brain (Fig. 8.32/9); after traversing the ventral portion of the pons, they reappear on the surface as the pyramids of the medulla oblongata (Fig. 8.19/17). Three fiber groups may be distinguished within the system: corticospinal fibers continue through the medulla oblongata into the spinal cord; corticobulbar fibers peel off at appropriate levels of the brainstem to reach lower motor neurons in motor nuclei of contralateral cranial nerves; and corticopontine fibers pass to various nuclei in the pons (Fig. 8.49/a-c).
Certain of the corticospinal fibers decussate within the medulla oblongata, and the others continue directly into the cord and decussate only when close to their terminations. The fibers with a medullary decussation form a lateral corticospinal tract within the lateral funiculus; those that continue uncrossed constitute a ventral corticospinal tract within the ventral funiculus (Fig. 8.18/3 and 10). The fibers of both tracts finally project on ventral gray matter motor neurons of the side contralateral to the fibers' origin in the cortex.
In domestic species, as in the generality of mammals, a short interneuron is often interposed between the upper and lower motor neuron; in primates, corticospinal fibers synapse directly with lower motor neurons in the spinal cord.There are other differences among species. In primates and carnivores, pyramidal fibers reach all levels of the cord; in the dog about 50% terminate in cervical segments, 20% in thoracic segments, and 30% in lumbosacrocaudal segments. In contrast, the pyramidal system of ungulates appears to have terminated completely by the level of origin of the brachial plexus (Fig. 8.50), although some corticospinal axons appear to travel the length of the spinal cord within the dorsal funiculus—the route, incidentally, that represents most of the corticospinal system in rodents. The proportion of fibers that decussate within the medulla oblongata also varies: about 50% do so in ungulates, 75% in primates, and all, or almost all, in the dog and cat.
The corticopontine fibers synapse on neurons in the nuclei of the ventral pons (Fig. 8.51/1); the axons of these second-order neurons then decussate and pass within the transverse fibers of the pons to enter the cerebellum through the middle cerebellar peduncle. Further successive synapses occur within the cerebellar cortex and then within the deep nuclei of the cerebellum (Fig. 8.51/8). From these cerebellar nuclei, axons travel back to the cerebral cortex via a relay through ventral thalamic nuclei (Fig. 8.51/11). This arrangement constitutes a pyramidal feedback system.
The Extrapyramidal System
The extrapyramidal motor system encompasses all brain areas involved in regulating motor functions that are not included within the pyramidal system. It is more complicated and involves various multisynaptic pathways that relay within a series of nuclei dispersed through the brain, from the telencephalon to the medulla oblongata. Some of these nuclei are large, grossly visible structures; others are small or diffuse, constituting a descending reticular system within the reticular formation of the brainstem.
Tracts originating in the tectum of the midbrain and in the lateral vestibular nucleus in the medulla are dealt with in the discussion of visual and vestibular pathways (p. 284).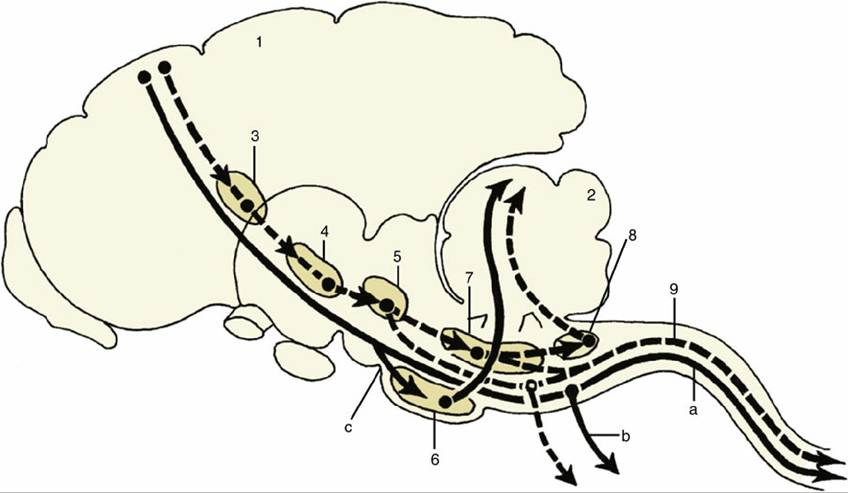
FIG. 8.49 Relay diagram of the pyramidal (solid lines and arrows) and the extrapyramidal (interrupted lines and arrows) systems. 1, Motor cortex; 2, cerebellum; 3, basal nuclei; 4, substantia nigra (mesencephalon); 5, red nucleus (mesencephalon); 6, pontine nuclei (metencephalon); 7, reticular formation; 8, olivary nucleus; 9, rubrospinal tract; a, corticospinal fibers; b, corticobulbar fibers; c, corticopontine fibers.
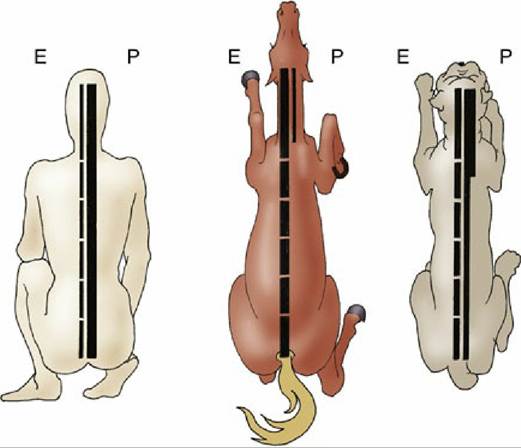
FIG. 8.50 Comparison of the pyramidal (P) and extrapyramidal (E) systems of the human, horse, and dog. The multisynaptic composition of the extrapyramidal system is indicated by the interruptions in this column; the width of a column is an indication of its relative importance.
The extrapyramidal system in part originates from regions of the cerebral cortex, including the primary motor area. Activity from these regions is relayed through a series of brain structures referred to collectively as the basal nuclei, which include the caudate nucleus, subthalamic nuclei, and substantia nigra, before returning via the thalamus to the cortex, thus forming a subcortical feedback loop. Other extrapyramidal output from the cortex reaches the red nucleus of the mesencephalon, the reticular formation, and the olive in the medulla oblongata (see Fig. 8.49). Only the red nucleus and the reticular formation contain neurons that project directly (or via interneurons) on the lower motor neurons of the brainstem and spinal cord.
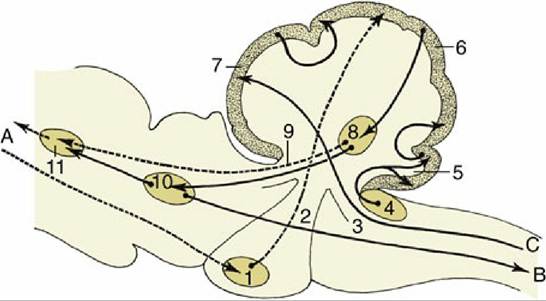
FIG. 8.51 Some important fiber connections of the cerebellum. The connections with the neocortex are represented by broken lines.
A, Tracts to and from the neocortex; B, tracts to the motor column of the spinal cord (extrapyramidal); C, proprioceptive tracts; 1, pontine nuclei; 2, middle cerebellar peduncle; 3, caudal cerebellar peduncle; 4, cochlear nuclei; 5, flocculonodular lobe of the cerebellum; 6, neocerebellum; 7, rostral cerebellar lobe; 8, cerebellar nuclei; 9, rostral cerebellar peduncle; 10, red nucleus; 11, thalamic nuclei.The axons from neurons in the red nucleus in the midbrain decussate immediately before descending through the ventrolateral part of the medulla oblongata to constitute a discrete rubrospinal tract bordering on the lateral corticospinal tract within the lateral funiculus of the cord (Fig. 8.18/4). This tract reaches the most caudal part of the cord, projecting en route on lower motor neurons in the ventral gray matter. This is an important tract in carnivores and is the best developed of all motor pathways in ungulates (see Fig. 8.50). It serves as a modulator of neural circuits that are located within the spinal cord gray matter.
The reticulospinal system is divided between well-defined dorsal and ventral tracts located within the lateral funiculus and a third (pontine reticulospinal) tract within the ventral funiculus (Fig. 8.18/II and III).
The activities of the various nuclei and connecting tracts of the extrapyramidal system are closely coordinated and so finely balanced that damage to any part may seriously impair the animal's ability to maintain posture or to execute intended movements. Different parts of the system play different roles: some are facilitatory, others inhibitory, and yet others facilitatory through removal of inhibitory influences. The numerous feedback circuits associated with the extrapyramidal system maintain the necessary balance between these facilitatory and inhibitory influences.
Cerebellar Function
The cerebellum does not itself initiate movement. Instead, it serves an essential regulatory function by ensuring that movements are executed as intended. To this end, the cerebellum receives a continuous stream of information from proprioceptors in the periphery via the subconscious proprioceptive pathways, from the vestibular apparatus via the vestibular nuclei, and from the pyramidal and extrapyramidal motor pathways via relays within the olivary nuclear complex (see Fig. 8.51). Thus, the cerebellum receives information about the current position and movements of the limb segments and of the head (proprioception and vestibular input) as well as information about the motor commands that have been sent to lower motor neurons controlling the limbs and head (pyramidal and extrapyramidal input). The cerebellum compares both inputs and sends any necessary corrective actions back to pyramidal and extrapyramidal circuits. The most important output from the cerebellum runs from the cerebellum to the thalamic nuclei and thence to the motor cortex and basal nuclei; other pathways travel to the contralateral red nuclei, the reticular formation, and the vestibular nuclei (for the coordination of vestibular reflexes).