SOME ASPECTS OF DEVELOPMENT
Only a few points need be raised to supplement the general account given in Chapter 5.
Most unusually, ovulation in cattle does not occur until some hours after the end of estrus.
Cleavage commences in the uterine tube, where fertilized ova are detained for several days before being released by the isthmus into the horn of the uterus. The small, spherical blastocyst that is first formed undergoes very rapid elongation from about the 13th day, first extending as a threadlike structure through the entire length of that horn and then, by about the 18th day, passing through the body to invade the contralateral horn. In this way, a single embryo takes maximal advantage of the endometrium available for its support. When twins are present, each claims one horn, and because both usually derive from the same ovary, transuterine migration seems to be readily accomplished. Contact between the two chorionic sacs is inevitable and results in fusion and anastomoses of the twin sets of vessels (with potentially unfortunate results; see Figure 29-28).The account of the development of the embryonic membranes and establishment of the cotyledonary placenta (see Figure 29-17) already given requires no amplification.
The placenta is a barrier to the intrauterine exchange of immune bodies in utero in ruminant species, and the newborn relies on colostrum for its early immunological protection.
Although the incidence of twin pregnancy in cattle is not high (1% to 4% according to breed), twinning has attracted much attention because of the virtual certainty that the female partner of a male calf will exhibit intersex characteristics. The masculinization of the female, the so-called freemartin, is due to exchange between the two circulations. It was long thought that exposure to androgens was the causal factor, but this is now believed to be of little importance.
In the prevailing view, what is significant is the transfer of antimullerian hormone (causing regression of the mullerian ducts) and descending (causing gubernacular outgrowth) and the exchange of cells between the two embryos, which are in fact chimeras (see Figure 29-18).Support for the last point is obtained from the fact that most cattle twins, presumably those that shared a common placental circulation, accept grafts of their partner’s skin in adult life, which indicates that cellular
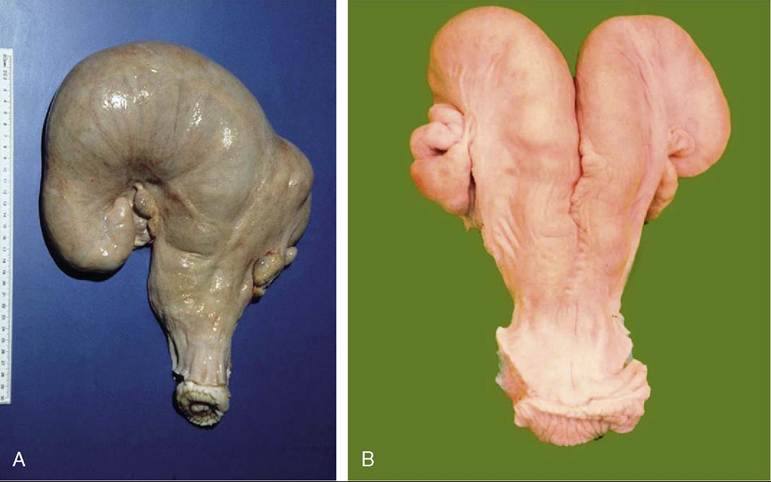
Figure 29-26 Changes in the uterus. A, Pregnancy involves mainly one horn, resulting in enormous asymmetry of the uterus. B, After pregnancy, the uterus returns to its former state; some asymmetry usually remains.
exchange had taken place when they were immunologically tolerant.
Twins and triplets are of course common in sheep and goats. The incidence varies with the breed and reflects the clemency or severity of the environment in which that breed evolved.
It is often convenient to be able to estimate the age of an aborted fetus in the field. There are many tables relating various measurements to age, but all suffer from the disadvantage of recording average values for parameters that vary considerably with breed, nutritional status, and other factors. One guide, easily memorized, allows 1 cm crown-rump length for each of the first 12 weeks’ gestation and 2.5 cm for each week thereafter. Except with the youngest embryos, it is rarely more than 2 weeks off, and greater accuracy is hardly to be expected of any rule-of-thumb method.
Qualitative methods that consider the external and internal anatomy are more accurate, but information on these matters must be sought elsewhere.
A few of the most obvious features are given in Tables 29-1 and 29-2.
Maturity in the sense of the capacity to make the integrated physiological responses necessary for survival outside the uterus is not achieved until late in gestation. In lambs the mortality is 100% in those delivered at 135 days and is still very high in those delivered at 140 days. Unfortunately, reliable information on these matters for cattle is not readily available.