Structure and Function of Blood Vessels
Microscopic Structure of
Blood Vessels
A single layer of endothelium, a simple squamous epithelium, lines all vessels. Capillaries are essentially endothelial tubes (Fig. 18-7), but they differ notably among themselves in cell-to- cell junctions and the structure of individual endothelial cells.
Tight junctions connect the endothelial cells of some capillaries, and this forms a relatively impermeable barrier to nonlipid-soluble particles. other capillaries appear to be quite porous because of openings in the endothelial cells themselves, and these are found where high rates of capillary exchange occur.A wall with three layers, or tunics, characterizes all types of veins and arteries. From innermost to outermost, these layers are the tunica interna (intima), tunica media, and tunica externa (adventitia) (Fig. 18-7). The tunica interna consists of the lining endothelium, a subendothelial layer primarily consisting of connective tissue, and in some cases, an internal elastic membrane. The internal elastic membrane consists of elastin and is especially prominent in large arteries.
The tunica media of all veins and arteries contains smooth muscle. Arteries have a much thicker tunica media than veins of a corresponding size, and this characteristic can be used to differentiate between the two types of vessels.
Elastic arteries are a special type whose tunica media contains concentric elastic laminae. The elastic properties of this type of artery permit vascular expansion and contraction during the various phases of the cardiac cycle. The aorta is an elastic artery that expands to increase its volume during ventricular systole and then rebounds during ventricular diastole to maintain relatively high arterial blood pressure. Arteries whose tunicae mediae consist primarily of smooth muscle are muscular arteries. An external elastic membrane is the outermost layer of the tunica media, but it is often difficult to visualize by light microscopy.
Arterioles, the smallest of arteries, are found where arteries empty into a branching capillary network.some veins, and especially those in limbs below the level of the heart, have flaplike valves that consist of folds of the tunica interna. The fold forms a cuplike pocket with the free edge of the fold directed toward the heart (Fig. 187). These one-way valves promote the flow of venous blood toward the heart when the vein is compressed.
Function of Blood Vessels
The arterial side of the circulation provides a ready supply of blood under relatively high hydrostatic pressure. Because of their relatively thick walls, arteries are not very compliant (i.e., do not distend easily with increases in pressure), so arterial pressure remains high as the heart pumps blood into the arteries. The arterioles at the end of the branching arterial network function as on-off valves to regulate the rate of blood flow from the arteries into capillary networks. Sympathetic vasoconstrictor nerves innervate the smooth muscle in the wall of most arterioles, and this is one mechanism by which blood flow is regulated. However, the degree of constriction of arteriolar smooth muscle is also subject to regulation by a large number of vasoactive agents, some locally produced (paracrine) and some that arrive via the systemic circulation. The paracrine agents (e.g., lactic acid, carbon dioxide, and adenosine) are typically vasodilators; they increase blood flow to the local area. Production of these agents increases when metabolism
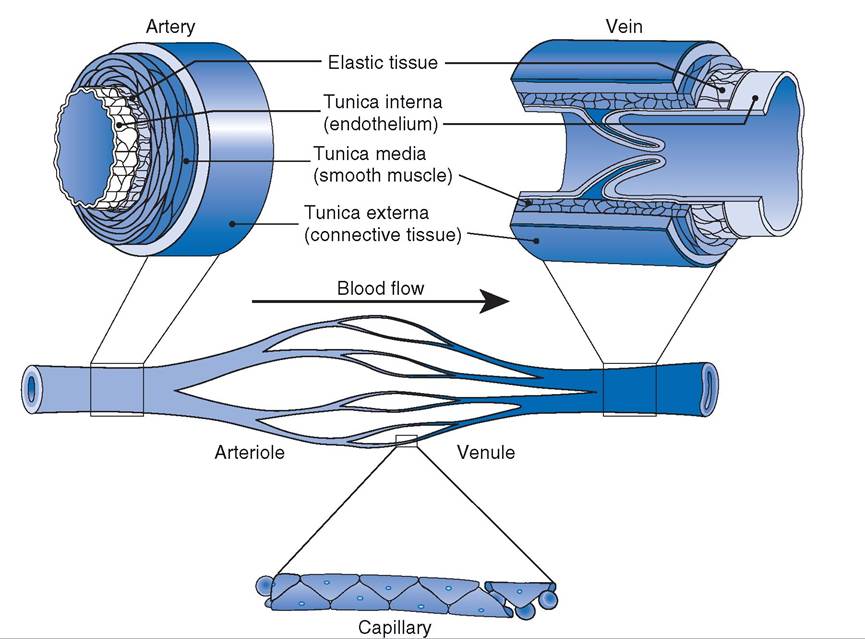
Figure 18-7. Section of small blood vessels showing the structure of their walls. A venous valve is also shown. (Reprinted with permission of Wolters Kluwer from Cohen B.J. and Wood D.L. MemmlerS The Human Body in Health and Disease. 9th ed. Baltimore: Lippincott Williams & Wilkins, 2000.)
by cells in the local area increases, providing a means to match metabolic rate with blood flow.
That is, an increase in metabolism brings about an increase in blood flow. The process by which local mechanisms regulate local blood flow is autoregulation.Capillaries are the site of exchange between blood and the interstitial fluid that surrounds all cells. in most cases, this exchange is by simple diffusion (i.e., substances move down their concentration gradients). Gases (oxygen and carbon dioxide) and other lipid-soluble substances freely diffuse though capillary walls, but substances that are not lipid soluble, such as glucose, must diffuse through pores in the capillary wall. Exchange by diffusion does not necessarily require the movement of fluid between the capillary and the interstitial space. oxygen, for example, can diffuse down its concentration gradient from the plasma to metabolizing cells as blood flows in a capillary past the cells. As stated earlier, the rate of capillary exchange is primarily governed by the rate of blood flow into the capillaries. in resting tissues, blood flow occurs only through a small percentage of the total capillaries at any one time. As metabolism and blood flow increase, the percentage of capillaries being perfused increases.
Typically, there is a small net loss of fluid from the plasma as it flows through most capillary networks. This fluid is recovered via the lymphatics and ultimately returned to the blood where lymphatics enter large veins near the heart. A small number of plasma proteins are similarly lost from capillaries and returned via the lymph.
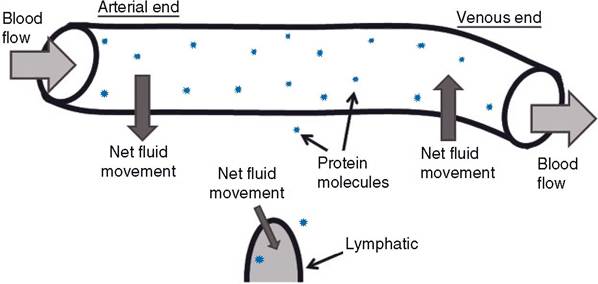
Figure 18-8. Microvascular fluid movements. Arrows indicate direction of net fluid movement. Note difference in relative amounts of protein molecules in plasma and interstitial fluid. Interstitial fluid with protein moves into lymphatics to form lymph.
The forces that govern fluid movement at the capillary level are important clinically in that imbalances in these forces contribute to edema, an abnormal amount or collection of fluid in the interstitial space.
The primary factor that forces fluid out of a capillary into the interstitial space is the blood pressure in the capillary. The primary force that tends to keep fluid in capillaries is the effective osmotic force (pressure) generated by plasma proteins, primarily albumin (Fig. 18-8). This pressure is also termed oncotic pressure. Plasma proteins generate an effective osmotic force because the protein concentration in the interstitial fluid is much lower than that of plasma. At the arterial end of a capillary, the blood pressure is higher than the oncotic pressure, so some fluid is lost from the capillary, while at the venous end of a capillary the oncotic pressure is higher, so some fluid moves into the capillary (Fig. 18-8). A slight imbalance between fluid loss and fluid gain by the capillaries gives rise to a net loss and provides fluid for lymph formation (Fig. 18-8).Like arteries, veins have smooth muscle in their walls, but the walls of veins are much thinner and more compliant. The compliance of venous vessels permits relatively large changes in the volume of blood in the veins with minimal changes in venous blood pressure. Thus, the venous side of the circulation functions as a low-pressure reservoir of blood. Constriction of venous smooth muscle promotes an increase in blood flowing back to the heart and an increase in cardiac filling pressure. This contributes to increases in cardiac output and the ability to perfuse capillaries for exchange.