The Cell Membrane
Structure of the Membrane
The outer cell membrane, also known as plasma membrane, and other membranes within the cell primarily consist of phospholipids, proteins, and cholesterol.
According to the fluid mosaic model, the phospholipids are arranged with their polar (hydrophilic) ends facing the protein layers, while their nonpolar (hydrophobic) ends face each other in the center of the membrane (Fig. 2-8). Because of the hydrophobic properties of the phospholipids, the membrane forms a water-impermeable barrier that separates the interior of the cell from the ECF. This barrier protects the cell by preventing the simple diffusion of water and water-soluble particles. Plasma membranes contain varying amounts of cholesterol, which is found between the phospholipid molecules and adjusts the fluidity and flexibility of the membrane.The protein composition of the outer cell membrane is extremely variable among different types of cells, and this variation has a great deal of influence on functional differences between cells. some membrane proteins are firmly inserted into the membrane among the phospholipids (Fig. 2-8). These are integral proteins; they may be arranged so that they extend completely across the membrane. Proteins that are exposed to both the cytosol inside
Figure 2-8. Fluid mosaic model of the cell membrane and proteins associated with the membrane.
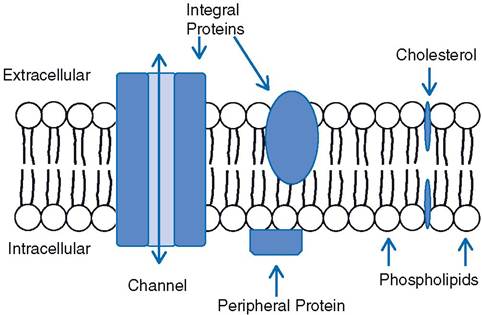
the cell and the extracellular fluid surrounding the cell are transmembrane proteins (Fig. 2-8). Many transmembrane proteins are involved in transport processes to move substances across the cell membrane. in Figure 2-8 a transmembrane protein forms a channel to permit passage across the cell membrane. Proteins may also be found bound to the surface of the membrane, and such proteins are classified as peripheral proteins (Fig.
2-8). Most of these are on the cytosolic surface of the membrane, and they are often found bound to an integral protein.The functions of cell membrane proteins include (1) transport of substances across the membrane; (2) provision of a site of binding for substances found in extracellular fluid, such as hormones; (3) contribution to the formation of cell-to-cell junctions; (4) provision of enzymes with active sites facing either the cytosol or the extracellular fluid; and (5) identification or recognition of cell type or cell origin by other cells. These functions are discussed in more detail later in this chapter.
structural modifications of the cell membrane occur largely on the free surface of cells (a surface not adjacent to any other cells). These modifications usually increase the cell surface and presumably function in absorption or secretion. The striated or brush border seen in light microscopy appears in electron micrographs to consist of uniform fingerlike projections called microvilli. Less regular projections, called stereocilia, are irregular branched extensions of the cell cytoplasm that are not motile.
Motile cilia (kinocilia) are complex elongated, fingerlike projections from cell surfaces found in areas where material is moved past the surface, as in the linings of the trachea and the uterine tubes. Each cilium is associated with a basal body that resembles a centriole normally seen in the cytoplasm of all cells.
Intercellular Contact and Adhesion
All vertebrates develop from division of a single cell, the fertilized egg. Unicellular animals also develop by division of a single parent cell. When the parent cell of a unicellular animal divides, the resulting daughter cells each go their own separate way, but the daughter cells of the fertilized ovum of a multicellular animal stay together and eventually differentiate into cells making up different tissues.
The ability of multiple individual cells to remain together and function as a tissue or organ depends on local modifications of the outer cell membrane.
in some cases, these modifications simply physically connect one cell to another. in other cases, these modifications connect cells and form a passageway for exchanges between them. The cell membrane modifications entail focal accumulations of specific membrane proteins termed cell adhesion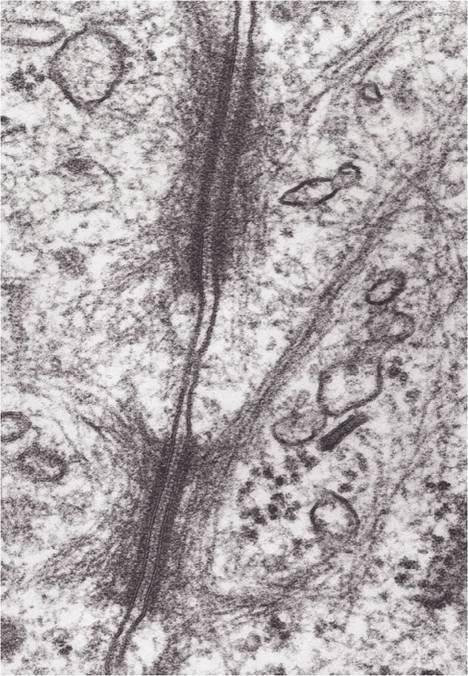
Figure 2-9. Two desmosomes connecting the membranes of two cells. (Reprinted with permission of Wiley-Blackwell from Dell- mann, H. D. and Eurell J. Textbook of Veterinary Histology. 5th ed. Baltimore: Lippincott Williams & Wilkins, 1998.)
molecules. The areas of cell membranes involved in intercellular contact and adhesion were named when they were initially examined by light and electron microscopy.
In electron micrographs, desmosomes appear as local thickenings of adjacent plasma membranes, with tiny fibrils radiating from the thickening into the cytoplasm of the cell (Fig. 2-9). Desmosomes tightly bind adjacent cells by the interactions between membrane proteins extending out from the surface of the cell membranes. They are seen as a single site or as a more extensive beltlike circle surrounding cells.
Tight junctions appear as an area or zone where the plasma membranes of two adjacent cells immediately adhere to each other. These are often found just below the free surface of epithelial cells. Each tight junction passes completely around the periphery of the cell at the same level. Tight junctions restrict the movement of water or dissolved materials into the space between adjacent cells.
Gap junctions are formed by membrane proteins that extend between adjacent cells to form a passage for exchange of small molecules and ions (Fig. 2-10). The exchange of ions permits one cell to affect the electrical activity of the adjacent cell. These types of exchanges have special functional importance in cardiac muscle and certain types of smooth muscle in the gastrointestinal tract.
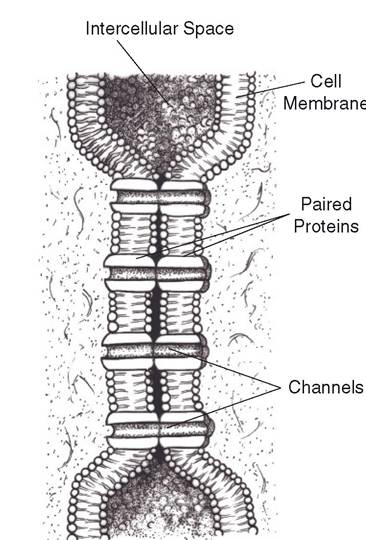
Figure 2-10. Two cell membranes connected at a gap junction by proteins that extend between them.