The Collecting Duct Reabsorbs Sodium Chloride and Can Secrete or Reabsorb Potassium
The collecting duct system begins with the connecting segment, which is a transition region from the distal convoluted tubule to the initial collecting tubule. Depending on the species, the connecting segment contains several distinct epithelial cell types, including distal convoluted tubule cells, connecting segment cells, intercalated cells, and principal cells.
Each of these structurally distinct cell types has specific physiological functions.The initial collecting tubules converge and empty into the cortical collecting duct, which descends through the cortex and medulla to the papillary tip, where the tubule fluid (urine) discharges into the renal pelvis. Phroughout most of the collecting duct system, two main cell types exist: the intercalated cell, which has many intracytoplasmic vesicles and mitochondria, and the principal cell, which has fewer intracytoplasmic vesicles and mitochondria but extensive basolateral plasma membrane infoldings (Figure 42-12). The principal cell is the major cell type in the initial collecting duct, the cortical collecting duct, and the outer medullary collecting duct, accounting for approximately two thirds of the cells in most regions. The intercalated cell accounts for the remainder of the cortical and outer medullary collecting duct cells, and in some species (rat, mouse, and human, at least) it persists even in the inner medullary collecting duct.
The principal cell reabsorbs NaCI in the collecting duct. Its extensive basolateral plasma membrane infoldings contain NaNK4-ATPase. As in other tubule segments, Na* is actively transported by this pump into the interstitial fluid, which establishes an electrochemical gradient for Na’ uptake from the tubule fluid through Na4 channels in the apical plasma
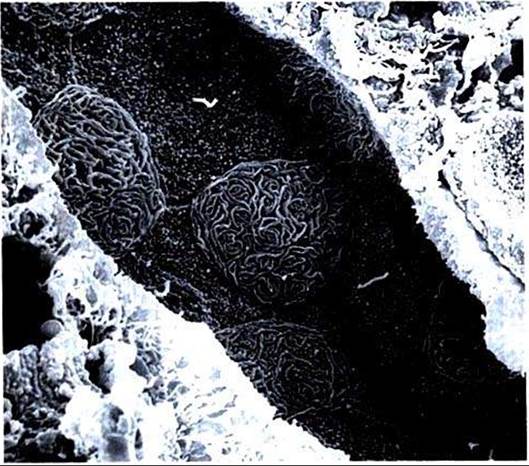
FIGURE 42-12 Scanning electron micrograph of outer medullary collecting duct in the rat, viewed from the luminal surface.Two cell types are evident: the principal cell, with short, small projections over the apical surface and a single central cilium, and the intercalated cell, with extensive, complex membrane folds (microplicae) over the apical surface. (Magnification ?2000.)
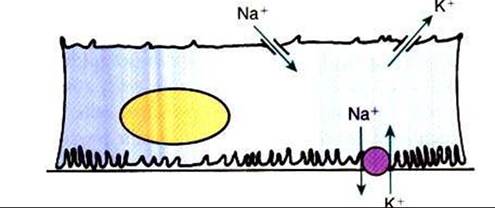
FIGURE 42-13 Schematic illustration of transport in the principal cell of the collecting duct.
The extensive basal plasma membrane infoldings contain large amounts of Na*,K+-ATPase. Active transport of Na* by this pump drives passive diffusion of Na* from the tubule lumen into the cell through a Na*-selective channel in the apical plasma membrane. A K*-selective channel in the apical plasma membrane provides a route for passive diffusion of intracellular K* into the tubule fluid.The hormone aldosterone enhances Na*,K+-ATPase activity and increases the Na* and K* permeability of the apical plasma membrane, thus enhancing Na* reabsorption and K* secretion in this segment.membrane. The resulting lumen-negative electrical potential drives CΓ absorption through the paracelkιlar pathway. At the same time, K1 is pumped actively from the interstitial fluid into the cell by Na+,K*-ATPase, raising the intracellular K+ concentration above that of the interstitial fluid and the tubule fluid. Intracellular K, exits the cell down the chemical gradient through K* channels present in the apical and baso- Iateral plasma membranes. Under normal circumstances, however, net K* secretion occurs for two reasons: (I) the apical K+ channel is more permeable than the basolateral K+ channel, and (2) the lumen-negative electrical potential favors K+ secretion (Figure 42-13).
The collecting duct can also reabsorb K+, and the intercalated cell appears to be responsible for this function. Potassium ions are actively transported from the cytoplasm across the apical plasma membrane of the intercalated cell in exchange for hydrogen ions by an Hf,K+-ATPase pump similar to that in the parietal cell of the stomach. This pump also contributes to the acidification of the urine (see Chapter 44).