THE DEFERENT DUCT
The deferent duct is undulating where it emerges but gradually straightens when followed toward the abdomen (Figure 5-42). It first runs medial to the epididymis as it heads toward the testicular vessels that form the bulkier components of the spermatic cord.
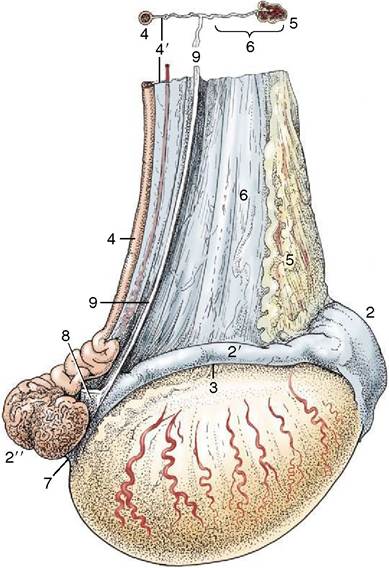
Figure 5—41 Lateral view of the right testis of a stallion. 1, Testis; 2, head of epididymis; 2', body of epididymis; 2'', tail of epididymis; 3, testicular bursa; 4, deferent duct; 4', meso- ductus deferens; 5, pampiniform plexus; 6, mesorchium; 7, proper ligament of testis; 8, ligament of tail of epididymis; 9, cut edge of fold connecting visceral and parietal layers of the vaginal tunic.
The constituents of the cord remain together as they pass through the inguinal canal but disperse at the vaginal ring (see Figure 5-36 and Figure 22-19). The duct here turns caudomedially to pass under the ureter before gaining the dorsal surface of the bladder (see Figure 5-36); it penetrates the prostate before finally entering the urethra a little way beyond the urethra’s origin from the bladder. The abdominal part continues to be supported by a peritoneal fold (mesoductus), which joins its contralateral partner to produce a horizontal genital fold above the bladder. The greater part of the duct is of uniform appearance and structure; its lumen is rather narrow in relation to the thick muscular wall. In most species the subterminal stretch lying on the bladder exhibits a fusiform enlargement, the ampulla of the deferent duct or ampullary gland (see Figure 5-51Z4). Although the term suggests a widening of the lumen, the thickening is mainly due to glandular proliferation in the wall of the duct, largely in the locally folded mucosa.
In most domestic mammals a second accessory gland grows from the duct close to its termination.
This, the vesicular gland, is described in a later section, but it may be noted in the meantime that the short, shared passage is known as the ejaculatory duct.The Vaginal Tunic and Spermatic Cord
The peritoneal process (vaginal tunic) that encloses the testis is an evagination of the lining of the abdomen through the inguinal canal. The narrow proximal part
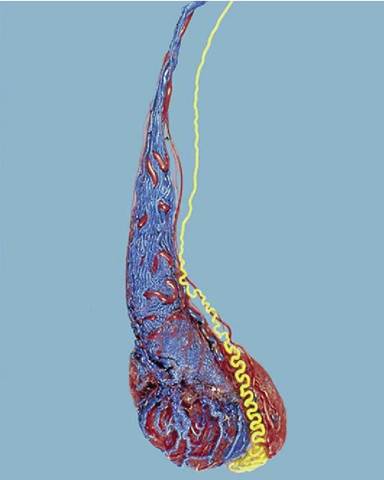
Figure 5-42 Corrosion cast (dog) of testicular artery (red), pampiniform plexus (blue), and deferent duct (yellow).
2
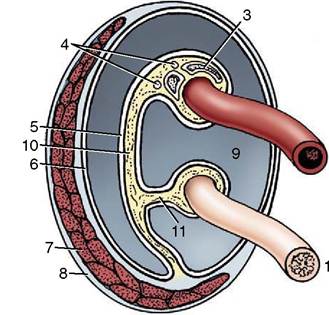
Figure 5-43 Transverse section of the spermatic cord and its immediate investments, schematic. 1, Deferent duct; 2, testicular artery (coiled); 3, pampiniform plexus; 4, testicular nerves and lymph vessels; 5, visceral layer of vaginal tunic; 6, parietal layer of vaginal tunic; 7, cremaster muscle; 8, external spermatic fascia; 9, vaginal cavity; 10, mesorchium; 11, mesoductus.
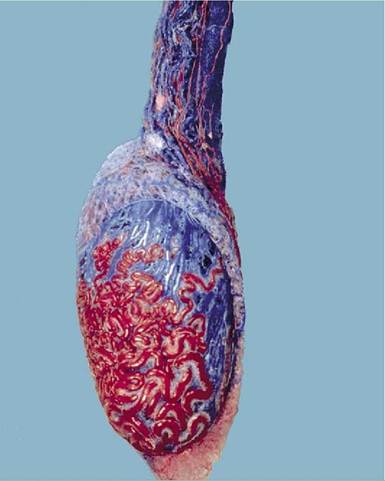
Figure 5-44 Corrosion cast of vessels within and on the testis and the pampiniform plexus (bull).
that surrounds the spermatic cord widens distally to form a flasklike expansion within the scrotum that encloses the testis and epididymis. The parietal and visceral layers of the tunic are connected by a fold that extends from the vaginal ring to the tail of the epididymis (see Figure 5-41). The cavity between the parietal and visceral layers (Figure 5-43/9) normally contains only a minute amount of serous fluid. It communicates with the peritoneal cavity of the abdomen through the vaginal ring, a narrow slitlike opening placed within the internal opening of the inguinal canal. Sometimes a loop of small intestine or another abdominal organ herniates into the peritoneal process through the vaginal ring; this complication is often encountered at castration.
It is worth mentioning that in human infants the neck of the peritoneal process usually becomes obliterated shortly after birth, which isolates the cavity about the testis.The spermatic cord varies in length and shape according to the position and orientation of the testis. It is shortest and most compact in those species in which the testis hangs vertically in the inguinal region. The bulk of the cord is provided by the testicular artery and veins, both remarkably modified. The artery branches from the abdominal aorta and first pursues a fairly direct course toward the vaginal ring, where the constituents of the spermatic cord are assembled. The more distal part is extraordinarily convoluted—one account describes no less than 7 m of artery packed within a 10-cm stretch of cord (Figures 5-44 and 5-45, A-B). These particular figures perhaps exaggerate the usual arrangement but serve to emphasize its extravagance. The testicular veins constitute a very elaborate close- meshed pampiniform plexus in which the contortions of the artery are embedded (Figure 5-45, B); the plexus ultimately reduces to a single vein that runs to the caudal vena cava. Arteriovenous anastomoses are present between the coiled testicular artery and its epididymal branches and the veins of the pampiniform plexus (Figure 5-46). A generous lymphatic drainage passes to lymph nodes placed about the bifurcation of the aorta. In some species a small lymph node is present near the inguinal canal. The lymph conveys a substantial fraction of the hormone production of the testis. The inconspicuous testicular nerves are of sympathetic origin.
The Scrotum
Variations in the location and form of the scrotum have been noted (see Figure 5-36). Externally, a median groove marks the division into right and left compartments; it often betrays a striking asymmetry of the testes. The lower part of the scrotum is molded on
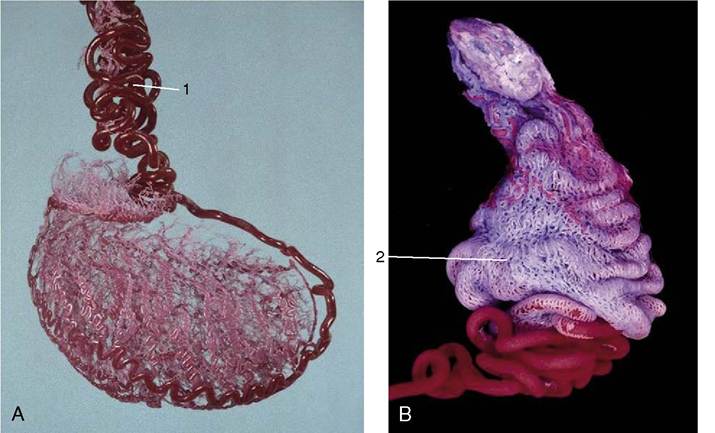
Figure 5-45 A and B, Vascularization of the equine testis. Observe the course of arterial branches on the testicular surface.
1, Testicular artery (red, becoming very tortuous as it approaches the testis); 2, part of the pampiniform plexus.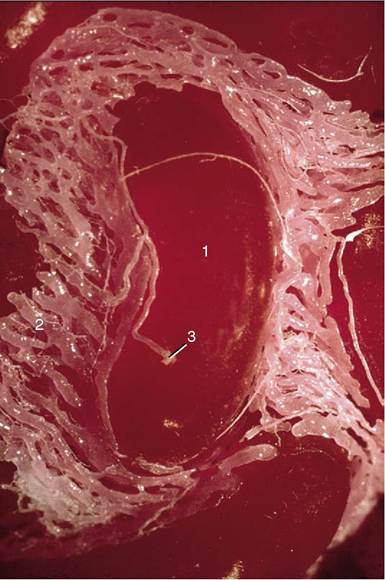
Figure 5-46 Corrosion cast of the testicular artery. 1, Coil of artery; 2, pampiniform plexus; 3, arteriovenous anastomosis (plexus filled via this anastomosis).
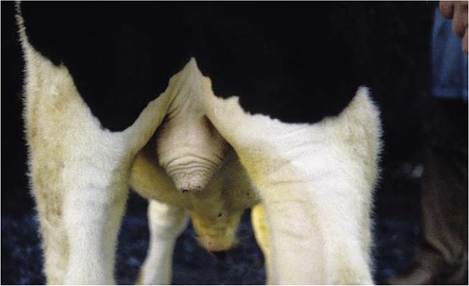
Figure 5-47 Scrotum of a bull. The musculature in tunica dartos is contracted causing elevation of the scrotum.
the testes and adjusts as their position varies with the ambient temperature (Figure 5-47).
The relatively thin scrotal skin is well provided with both sweat and sebaceous glands. It is sometimes rather bare, but this is not a constant feature; indeed, the scrotum is hidden by hair in the cat and densely covered by fleece in sheep of certain breeds. When bare, it is often pigmented. The scrotal skin adheres to a tough Abromuscular layer (tunica dartos), which also extends as a septum between the compartments that separately lodge the testes. Internal to the dartos, a (spermatic)
fascia is present that may be resolved into several layers, which are believed to correspond to the layers of the abdominal wall. The predominant layer is the external spermatic fascia, which can be clearly separated from the dartos (Figure 5-48). The loose intermediate stratum allows the vaginal tunic independent movement within the scrotal sac; in addition to its functional significance (see further on), this facilitates castration by the closed method (in which the testis is brought to the exterior within the vaginal tunic before the cord is severed proximally). The dense external spermatic fascia that supports the vaginal tunic also invests the cremaster, a slip of muscle that passes onto the cord on detachment from the caudal margin of the internal oblique muscle of the abdomen.
Testicular Function
In most wild mammals the breeding period is seasonal, and this is reflected by changes in the morphology and activity of the reproductive organs of both sexes.
Little of this seasonality remains among male domestic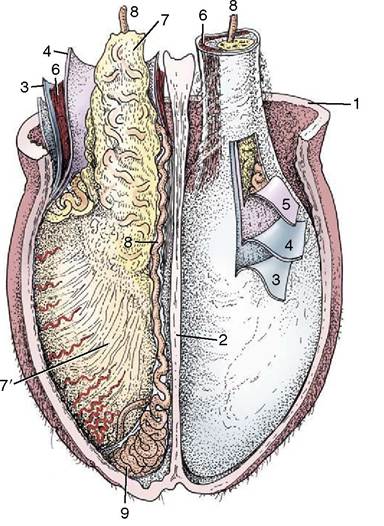
Figure 5-48 Cranial view of the opened scrotum of a bull; the investments of the testis have been partly dissected. 1, Scrotal skin and dartos; 2, scrotal septum; 3, external spermatic fascia; 4, parietal layer of vaginal tunic; 5, visceral layer (dissected from surface of testis); 6, cremaster muscle; 7, visceral layer of vaginal tunic covering structures in spermatic cord; 7, visceral layer on testis; 8, deferent duct; 9, tail of epididymis.
animals, in which the seminiferous epithelium is active throughout the year with at most only slight variation in sperm output. Although the process of spermatogenesis is not described, the reader is reminded that the serial cell divisions and maturation processes that constitute the cycle are not synchronous in every part of the seminiferous epithelium. Instead, adjacent segments show successive stages so that a “lucky” longitudinal section of a tubule displays the different stages of the process occurring as a wave spreading along its length (see Figure 5-39).
The process of spermatogenesis is influenced by temperature, and as already stated, it cannot proceed normally at the core temperature of the body. The seminiferous epithelium is damaged in testes that fail to descend into the scrotum (the “cryptorchid condition”), and these do not produce sperm. Similar changes are evident in testes that, having descended successfully, are later returned to the abdomen and, indeed, in scrotal testes that are overheated by an unusually thick covering of hair or fleece. Because the interstitial tissue is less susceptible to temperature, it follows that libido and potency may be normal in cryptorchid animals that are infertile.
Many factors help maintain the appropriate endotes- ticular temperature. The exposed position of the scrotum, the absence of fat within the scrotal fascia, and the intracapsular situation of large testicular vessels all favor heat loss by radiation (Figure 5-49); the generous supply of sweat glands allows additional loss through evaporation from the skin surface.
Perhaps more importantly, the extensive contact between the vessels within the cord precools the blood within the artery as this follows its winding course in relation to the venous plexus (see Figure 5-45). The opportunities for heat loss are such that the testicular temperature could be lowered excessively in colder climates. Countermeasures are available. Contraction of the tunica dartos, directly sensitive to temperature change, tightens and bunches the scrotum, thereby reducing the exposed surface and also drawing the testes toward the warmer trunk (see Figure 5-47). The testes may also be separately raised within the scrotum by contraction of the cremaster muscles, which pull on the vaginal tunics; being striated, these muscles react briskly to pull the testes away from potentially harmful stimuli.Castration of surplus male animals has long been practiced to make them more manageable or to promote particular carcass qualities. Modern husbandry, the effects of selective breeding, and changes in consumer requirements now make it possible to bring food animals to slaughter at earlier ages than before, and the necessity for and economic advantage of routine castration are beginning to be questioned. The direct influence of cas-
tration on the reproductive organs is considered in some detail for cattle, the species about which most is known, on page 719.