The Forebrain
The forebrain comprises the diencephalon and the paired cerebral hemispheres (telencephalon). The hemispheres overlap the dorsolateral aspects of the diencephalon, to which they are connected by extensive fiber tracts.
The Diencephalon
The diencephalon (there is no convenient alternative name) forms the most rostral part of the brainstem. Only its most ventral part, the hypothalamus, is visible on the external surface of the intact brain (see Fig. 8.19), but it is more extensively revealed in median section (see Fig. 8.21). The diencephalon has three parts: epithalamus, thalamus (including subthalamus), and hypothalamus, which develop in relation to the roof, walls, and floor of the third ventricle, respectively.
The epithalamus, the most dorsal part, comprises the pineal gland (epiphysis cerebri), habenular striae, habenulae, and habenular commissure (Fig. 8.30). The pineal gland (Fig. 8.30/6) is a small, median body projecting dorsally from the brainstem behind an evagination of the roof of the third ventricle that is composed only of pia and ependyma. The pineal gland plays an important role in sexual development and behavior; it is believed to be particularly concerned in the seasonal regulation of ovarian activity in response to changing day length. The pineal gland produces melatonin, which is important in circadian and seasonal rhythms (p. 206). The habenulae are nuclear complexes that develop within the most dorsal parts of wall of the third ventricle. They have important functions relating to emotion, motivation, and reward. The nuclei receive fibers (habenular stria) from the hippocampus and other parts of the telencephalon and send fibers to mesencephalic nuclei. The left and right habenular nuclei are interconnected via the habenular commissure (Fig. 8.30/5').
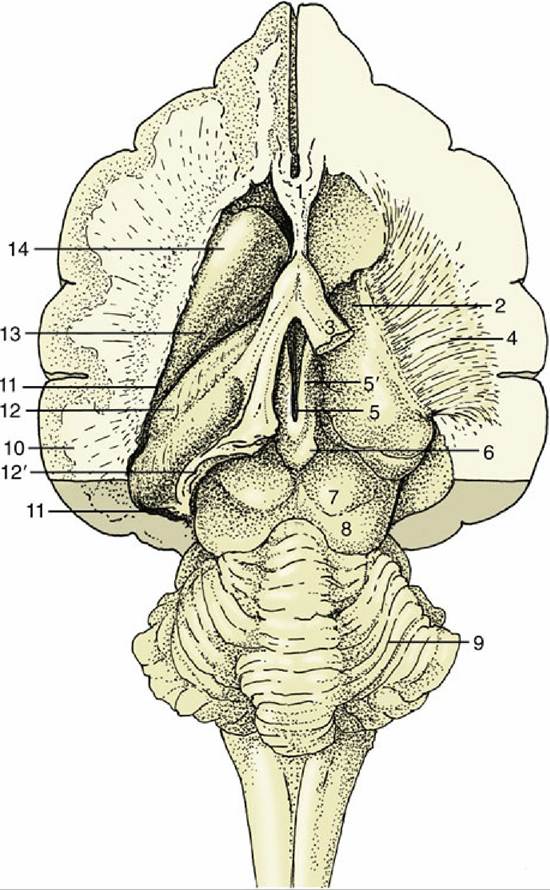
FIG.
8.30 Dorsal view of the canine brain. Part of the left hemisphere has been removed, opening the lateral ventricle. On the right, the hippocampus and basal nuclei have also been removed, exposing the thalamus and the internal capsule. 1, Septal nuclei; 2, dorsal surface of thalamus; 3, fornix (cut); 4, internal capsule; 5, dorsal part of third ventricle; 5', habenular nuclei (in roof of third ventricle); 6, epiphysis; 7, rostral colliculus; 8, caudal colliculus; 9, cerebellum; 10, cut lateral wall of hemisphere; 11, lumen of lateral ventricle; 12, hippocampus; 12', cut-edge of denticulate gyrus; 13, tail of caudate nucleus; 14, head of caudate nucleus.The thalamus is the largest component of the diencephalon. It develops within the lateral walls of the third ventricle, but in many species, including domestic species, the wall of each side of the thalamus expands medially into the ventricle to connect right and left sides. This, the intermediate mass or interthalamic adhesion, reduces the ventricle to an encircling annular space (Fig. 8.31/3). The relations of the thalamus are difficult to envisage because of its deep position and lack of separation from neighboring structures. It extends rostrally to the lamina terminalis grisea and to the midbrain caudally. Its dorsal surface contacts the fornix, a telencephalic structure, and the floor of the lateral ventricle; its ventral surface is continuous with the hypothalamus; and its lateral face is enclosed by an internal capsule of white matter containing axons ascending to and descending from the cerebral cortex (see Fig. 8.30).
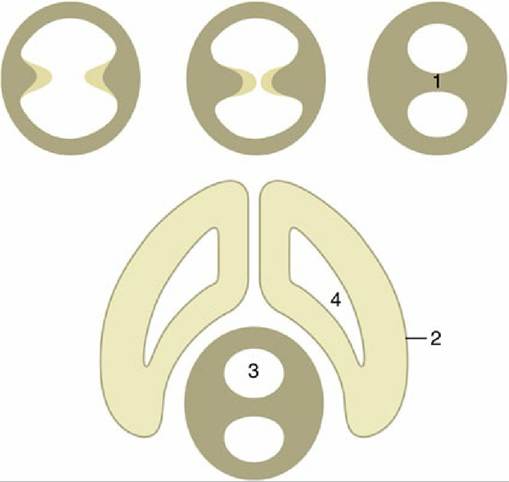
FIG. 8.31 Top left to right, The formation of the interthalamic adhesion (1) by median fusion of outgrowths of the lateral walls of the diencephalon. 2, Telencephalon; 3, third ventricle; 4, lateral ventricle.
The thalamus is composed of a very large number of nuclei named according to their topographic relationships to one another.
These nuclei have various specific functions and collectively form one of the most important relay and integration centers of the brainstem. Nuclei in the ventral thalamus receive information from most afferent systems (excluding the pathways concerned with olfaction) and also provide relays on feedback control systems of motor pathways (Fig. 8.32).The subthalamus contains the subthalamic and endopeduncular nuclei and the zona incerta. The subthalamic nucleus acts as a relay station for the extrapyramidal motor pathway, whereas the other nuclei serve as links between the limbic system and the somatic and visceral motor systems.
The metathalamus, the caudolateral part of the thalamus, comprises the lateral and medial geniculate bodies (Fig. 8.33/3 and 5), whose presence and position were noted in the description of the midbrain. The lateral geniculate body, although not conspicuous in itself, is joined by the optic tract, which sweeps caudodorsally over the surface of the thalamus. The medial geniculate body lies ventromedial to the lateral geniculate body and receives acoustic fibers, the brachium, via the caudal colliculus (p. 287). The nuclei within the lateral and medial geniculate bodies relay visual and acoustic information, respectively, to the cerebral cortex.
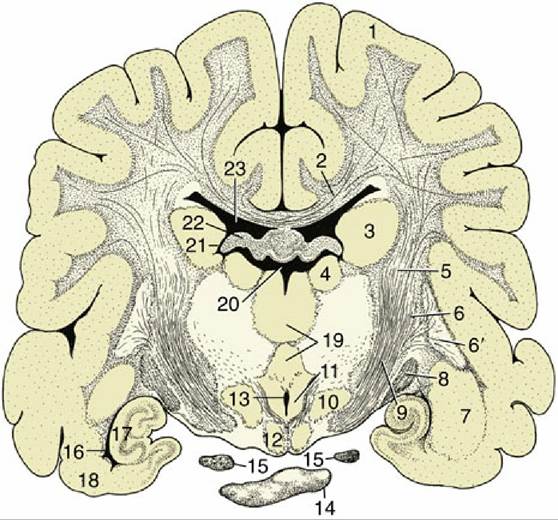
FIG. 8.32 Transverse section of the canine brain at the transition between the crus cerebri and internal capsule. 1, Cerebral hemisphere; 2, corpus callosum; 3, caudate nucleus; 4, thalamic nuclei; 5, internal capsule; 6 and 6', lentiform nucleus: 6, globus pallidus; 6', putamen; 7, amygdala; 8, optic tract; 9, crus cerebri; 10, hypothalamic nuclei; 11, mammillothalamic tract; 12, mammillary body; 13, ventral part of third ventricle; 14, hypophysis; 15, oculomotor nerve; 16, ventral part of lateral ventricle; 17, hippocampus; 18, piriform lobe; 19, interthalamic adhesion; 20, dorsal part of third ventricle; 21, interventricular foramen; 22, fornix; 23, lateral ventricle.
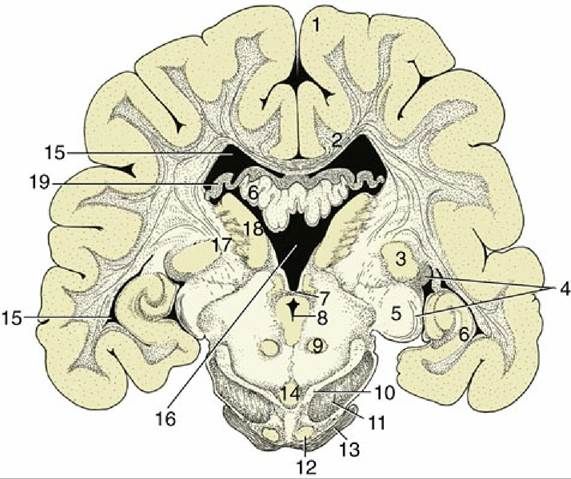
FIG. 8.33 Transverse section of the canine brain at the boundary between the mesencephalon and diencephalon. 1, Cerebral hemisphere; 2, corpus callosum; 3, lateral geniculate nucleus; 4, optic tract; 5, medial geniculate nucleus; 6, hippocampus; 7, caudal commissure; 8, mesencephalic aqueduct; 9, red nucleus; 10, substantia nigra; 11, crus cerebri; 12, rostral extension of pontine nuclei; 13, middle cerebellar peduncle; 14, interpeduncular nucleus; 15, lateral ventricle; 16, third ventricle; 17, internal capsule; 18, thalamic nuclei; 19, fornix.
The hypothalamus forms the ventralmost parts of the lateral walls of the third ventricle. It appears
on the external surface of the brain between the preoptic region (rostral to the optic chiasm) and the cerebral peduncles and interpeduncular fossa caudally (see Fig. 8.19). Its salient surface features are, first, the region known as the tuber cinereum, from which extends the stalk or infundibulum that suspends the hypophysis below the brain, and second, the rounded mammillary body (see Fig. 8.21) that receives information from the hippocampal complex and sends information to the thalamus via the mammillothalamic tract. The mammillary body is an important structure for memory. Internally, the hypothalamus contains a number of nuclei associated with the visceral nervous system as well as with hormonal and homeostatic regulation.
The hypophysis, or pituitary gland, is a dark solid body. It is located within a recess of the floor of the cranial cavity and is usually left behind when the brain is removed because the infundibulum, hollowed by a recess of the third ventricle, is easily torn across. The hypophysis is also held in place by a fold of dura mater (p. 295). The functions of the hypophysis are described elsewhere (p. 203).
The Telencephalon (Cerebrum)
The telencephalon consists of the paired hemispheres and the lamina terminalis grisea, the thin plate forming the rostral wall of the third ventricle with the vascular organ of the lamina terminalis (see Fig.
8.67/7). Because the hemispheres develop as outgrowths of the diencephalon, their walls and lumina (lateral ventricles) remain in direct continuity with the corresponding regions of the diencephalon. The adult hemispheres are semiovoid structures that form the largest part of the brain; their growth causes them to extend caudally over the brainstem to reach to within a short distance of the cerebellum. This growth brings them close together, and their flattened medial surfaces face toward each other across the narrow longitudinal fissure into which the falx cerebri, a connective tissue sheet, fits when the brain is in situ. The remainder of the outer wall is divided between convex dorsolateral and flattish ventral (basal) surfaces (see Figs. 8.23, 8.32, and 8.33).The walls of the hemispheres thicken unequally. Much of the medial wall of each hemisphere remains particularly thin, and in fetal life a part rolls inward, invaginating the pia mater and blood vessels covered by the ependymal lining into the ventricle, where it develops into the choroid plexus (p. 296) associated with this cavity. This structure produces the cerebrospinal fluid. The ventrolateral (striatal) part of the wall becomes much thickened when a number of large nuclei, the basal nuclei, develop within it. The alternation of these nuclei with the fiber tracts in which they are embedded lends this region a striated appearance when exposed by section (see Fig. 8.32); it is therefore appropriately known as the corpus striatum. The remainder of the wall is initially known as the pallium, but when it acquires an external covering of gray substance, by migration of cells from the ependyma during development, it is more frequently termed the cortex, although this term strictly designates only the outer gray substance.
Three regions of the pallium (or cortex) are distinguished on the basis of evolutionary history, structure, and function. The paleopallium initially served a purely olfactory function; it has retained this association in highly developed mammals.
The archipallium was also initially concerned with olfaction, but unlike the paleopallium, it has largely lost this association. The youngest part, the neopallium, made a very modest initial appearance in vertebrate history but has undergone a spectacular enlargement in mammals, in which it is both the largest and the functionally dominant part of the mammalian telencephalon. These parts are now described separately but in a different order for convenience. First, it may be helpful to dispose of the concept of a rhinencephalon ("smellbrain") of primary olfactory function. Although it is true that the telencephalon of lower vertebrates developed specifically in relation to this sense, many parts have since discarded their original function and acquired new roles. The term rhinencephalon therefore no longer describes the functions of these parts at all adequately, and because it is now used in many conflicting ways, there is little in favor of its retention.The Paleopallium
The paleopallium is confined to the basal part of the brain; it is separated from the neopallium by the rhinal sulcus (Fig. 8.34/4) on the lateral surface and, although less clearly, from the archipallium medially. Its rostral extremity is provided by an appendage, the olfactory bulb (Fig. 8.34/1), that fits into a recess of the ethmoid bone. The surface apposed to the bone is made shaggy by the entrance of the numerous filaments that together form the olfactory nerve (I); these arise from receptors within the nasal mucosa and pass through the many perforations in the cribriform plate of the ethmoid bone. In the olfactory bulb, the olfactory stimuli are conveyed to second-stage neurons.
The bulb is continued caudally by the common olfactory tract (see Fig. 8.19/2), which soon divides into medial and lateral divisions separated by a triangular area. The medial tract runs toward the medial aspect of the hemisphere (precommissural area), where the information is conveyed to third-stage neurons. Some of the continuing fibers terminate within certain cortical gyri; others pass through the narrow anterior commissure in the rostral wall of the third ventricle to reach the corresponding region of the opposite hemisphere. The lateral tract continues caudally to join the large piriform lobe (see Fig. 8.19/3), the most salient feature of the basal surface of the hemisphere; not all the fibers in this tract reach the piriform lobe; some synapse on deeper structures such as the amygdala.
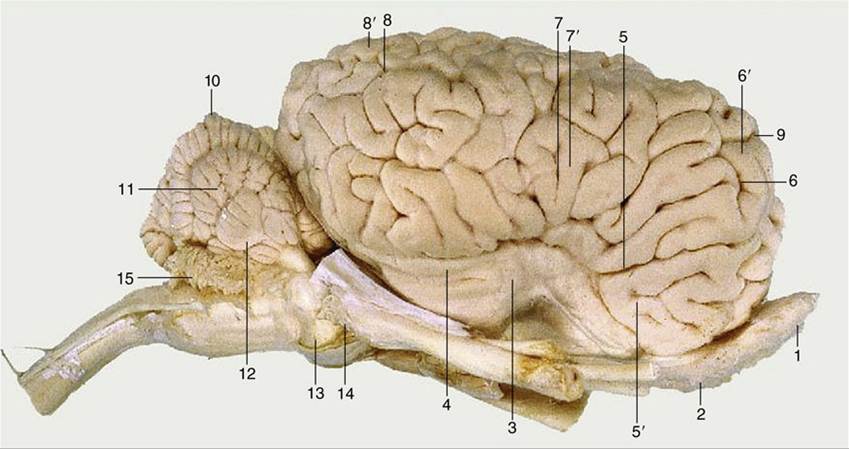
FIG. 8.34 Lateral view of the equine brain. 1, Olfactory bulb; 2, olfactory tract; 3, piriform lobe; 4, rhinal sulcus; 5, sylvian sulcus; 5', sylvian gyrus; 6, ectosylvian sulcus; 6', ectosylvian gyrus; 7, suprasylvian sulcus; 7', suprasylvian gyrus; 8, ectomarginal sulcus; 8', ectomarginal gyrus; 9, cruciate sulcus; 10, cerebellar vermis; 11, cerebellar hemisphere; 12, paraflocculus; 13, pons; 14, crus cerebri; 15, caudal medullary velum.
The Basal Nuclei
The basal nuclei are a group of large nuclei positioned dorsal to the paleopallium; they consist of the caudate nucleus and the putamen (which together are referred to as the striatum, or neostriatum) and the globus pallidus, or paleostriatum. The basal nuclei coordinate activity between the cerebral cortex and thalamus and play an important role in the initiation and regulation of movement. The caudate nucleus (Fig. 8.32/3) has the general form of a comma with a large head bulging into the floor of the main part of the lateral ventricle, a body following the caudal bend of the ventricle, and a tail close to the roof of ventral extension of the ventricle (Fig. 8.30/13 and 14). The caudate nucleus and the more laterally located putamen are functionally one structure, although anatomically they appear as two nuclei separated by a band of white matter that is composed of the axons of the internal capsule passing to and from the cerebral cortex. The globus pallidus, the other component of the basal nuclei, is also located laterally and, when combined with the putamen, is referred to as the lentiform nucleus (Fig. 8.32/6 and 6'). The lentiform nucleus is separated from the thalamus by the caudal portion of the internal capsule. The nucleus accumbens, the reward center, is located in the ventral part of the neostriatum.
The other components of the basal nuclei complex are the smaller amygdala (Fig. 8.32/7), located near the tail of the caudate nucleus, and the claustrum, which is interposed between the lentiform nucleus and neopallium. It is separated from these structures by other white matter tracts; the one on the lateral face of the claustrum is known as the external capsule.
The Neopallium
The neopallium makes up the majority of the telencephalon: all of the telencephalon that is visible in dorsal view and the bulk of that visible in lateral and medial views is neopallium. It is commonly referred to as the cerebral cortex. It is divided from the ventrally located paleopallium by the rhinal sulcus on the lateral side of the hemisphere (Fig. 8.20/4) and from the archipallium, described later, by the splenial sulcus medially (Fig. 8.21/4). In mammals of smaller size, rats and mice for example, the outer surface of the neopallium is smooth; however, in larger mammals, including domestic species, it displays a complicated arrangement of alternating ridges (gyri) and grooves (sulci) (see Fig. 8.23). Though it is tempting to regard the presence of more intricate gyri and deeper sulci as evidence of greater intelligence and increased capacity for complex responses, the immediate underlying cause appears to be physical. The ridges, which are mainly longitudinal, are produced by restraints imposed on the expanding telencephalic vesicle by the rigid corpus striatum and corpus callosum, and additional folding is necessary to maintain the relationship between volume (which increases by the cube) and cortical area (which increases by the square) in large brains.
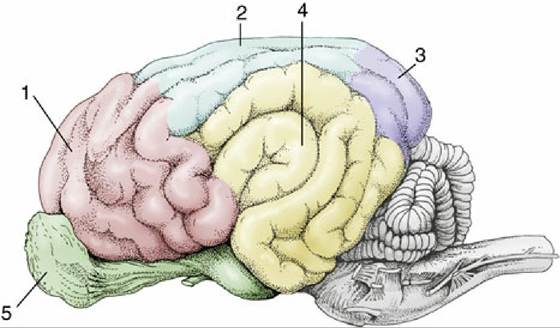
FIG. 8.35 Cortical lobes of the canine brain, lateral view. 1, Frontal lobe; 2, parietal lobe; 3, occipital lobe; 4, temporal lobe; 5, olfactory lobe.
The pattern of the gyri is reasonably constant within one species but differs among species. The features of greatest consistency include the cruciate sulcus, running transversely on the rostrodorsal aspect, and the sylvian sulcus on the lateral side. Although other features provide useful landmarks for the investigator seeking to establish the functional significance of particular cortical areas, the names of most are of little consequence to the student. A simpler, rather arbitrary division of more general utility distinguishes four regions or lobes named for their proximity to overlying bones; this division recognizes frontal, parietal, and occipital lobes in rostrocaudal sequence and a temporal lobe lying lateral to the last two. Only the frontal lobe is clearly demarcated because it is bounded caudally by the cruciate sulcus (see Figs. 8.23/14 and 8.35). Functional regions of the cerebral cortex have also been identified through experimentation. Thus we can locate the motor cortex, the sensory cortex, the auditory cortex and the visual cortex (Figs. 8.20B and 8.23B).
The concept of somatotopy is a characteristic of many structures in the brain and spinal cord but is expressed most highly in the cerebral cortex and to a lesser extent in the cerebellar cortex and so is discussed here. The fibers and cell bodies within many tracts and relaying nuclei and within areas of the cerebral and cerebellar cortices preserve very orderly point-to-point arrangements that reflect the topography of the parts of the body from which afferent impulses arise or to which efferent impulses are delivered. These do not always, or even usually, reproduce the true physical proportions but represent the parts of the body in relation to the densities of their innervation. The representations take the form of grotesque caricatures, sometimes known as homunculi—although animalcula would better fit veterinary anatomy—in which very sensitive, densely innervated body regions such as the lips and muzzle of the horse, or regions capable of very refined and accurate movements, such as the fingers of a human or the prehensile tail of a monkey, are of exaggerated size. The concept of somatotopy is of great importance in the consideration of the significance of pathologic lesions, in the conduct of neurosurgery, and in experimental stimulation.
The histologic structure of the neopallium is remarkably uniform, consisting of six superimposed strata that are densely populated by neurons and are separated by cell-free divisions. The neurons are broadly of two types: some more or less spherical (granular) neurons are provided with processes of very limited extent, and other, pyramidal, neurons have processes that range more distantly within the underlying white substance. The pyramidal neurons can be classified by the connection of their axon fibers into association neurons, commissural neurons, or projection neurons. Association fibers connect parts of the neopallium of the same hemisphere after passage directly below the cortex. Commissural fibers connect the two hemispheres, generally linking equivalent contralateral parts. They run over the roof of the lateral ventricle and mainly cross within the corpus callosum, the major telencephalic commissure that is shaped to form a rostral genu, middle trunk, and caudal splenium (Fig. 8.21/3). Descending projection fibers from the cortex connect with lower parts of the central nervous system; most converge on the internal capsule squeezed between the basal nuclei and thalamus (Figs. 8.36/7 and 8.37/1). In their courses to, from, and within the capsule the projection fibers are ordered according to their functional associations and somatotopic relationships.
The Archipallium
The part of the cortex known as the archipallium is concerned with the correlation of olfactory with other sensory information but has acquired additional functions in modern mammals. It forms part of the limbic system, which comprises the cingulate, supracallosal, and geniculate gyri; the hippocampal formation; and the dentate gyrus.
The archipallium is not a conspicuous feature of the telencephalon in domestic mammals. The enormous development of the neopallium has caused the archipallium to be displaced to the medial wall of the cerebral hemisphere; it is further reduced in prominence because a large part is rolled inward to lie on the floor of the lateral ventricle. The archipallium is topographically divided by the corpus callosum into a dorsal part that remains on the medial surface of the hemisphere named the cingulate and supracallosal gyri between the splenial sulcus and the corpus callosum (Fig. 8.21/8 and 8') and a ventral part composed of the inflected portion, usually known as the hippocampus (Fig. 8.38/2), located deep in the hemisphere. The archipallium is curved in conformity with the shape assumed by the expanding telencephalon and fits around the dorsal, caudal, and ventral aspects of the thalamus. This arrangement is difficult to envisage, and it is helpful to remember that the archipallium is interposed between the olfactory bulb and the hypothalamus. The pathway is thus bent into a hairpin loop by the expansion of the hemisphere (Fig. 8.39); the proximal limb extends, with a ventral concavity, caudally toward the apex of the loop, where a spiral twist sets the distal limb on a parallel returning course.
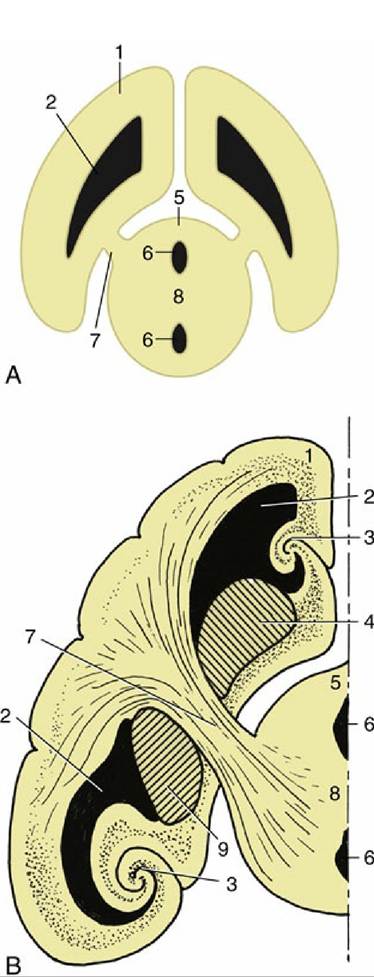
FIG. 8.36 (A) The connection between the cerebral hemisphere and diencephalon via the internal capsule (7). (B) The lateral ventricle, basal nuclei, and hippocampus form concentric arches over the internal capsule. 1, Cerebral hemisphere; 2, lateral ventricle; 3, hippocampus; 4, caudate nucleus; 5, diencephalon; 6, third ventricle; 7, internal capsule; 8, interthalamic adhesion; 9, globus pallidus and putamen.
The proximal limb is formed by the medial surface gyri; ventral to this run the longitudinal association fibers (cingulum) that originate in the septal area. The fibers of this multisynaptic pathway enter the caudal extremity of the hippocampus and form a covering for it. The fibers leaving the hippocampus run rostrally over its surface, gradually consolidating into a thick bundle, the fornix. The fornix lies directly below the corpus callosum at its start but then deviates ventrally as it passes rostrally; it curves around the rostral extremity of the thalamus to enter the hypothalamus, where it terminates within the mammillary body (Figs. 8.38 and 8.40). The right and left hippocampi are joined by the commissure of the fornix. There are thus three telencephalic commissures: the large corpus callosum of the neopallium, the anterior commissure of the paleopallium, and the commissure of the fornix (also known as the archipallial fornical commissure).
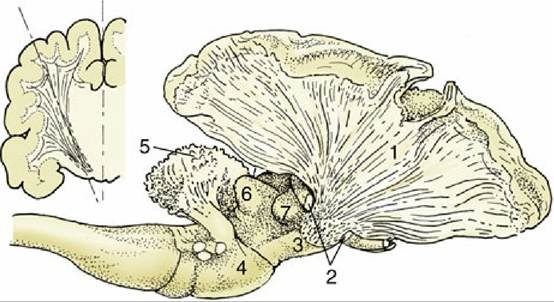
FIG. 8.37 The internal capsule in the canine brain. A part of the cerebral cortex and the cortex of the cerebellum have been removed. The resected part of the telencephalon is indicated in the inset. 1, Fibers of the internal capsule; 2, optic tract, partly removed; 3, crus cerebri; 4, pons; 5, corpus medullare of cerebellum; 6, caudal colliculus; 7, medial geniculate body.
When the fornix parts company with the corpus callosum, it remains connected to it by a thin vertically oriented septum that increases in depth toward its rostral end. This septum telencephali (pellucidum) forms part of the medial wall of the lateral ventricle (see Fig. 8.21/28). It is a bilateral structure, each sheet of which is separated from its neighbor by a narrow, completely enclosed cleft, and its ventrorostral part contains septal nuclei in which fibers from the medial olfactory tract terminate.