Processing of Somatic Afferent Information
The term somatic afferent is applied to those sensory pathways that receive and convey information from the wide array of peripheral receptors scattered throughout the skin and the deeper somatic tissues.
The term does not apply to the afferent pathways from the retina of the eye or the inner ear (referred to as special afferent pathways) or those from receptors in the viscera (visceral afferent pathways).The somatic afferent system transmits a variety of sensory modalities: touch, pressure, vibratory sensation, thermal sensation, pain, as well as kinesthetic or proprioceptive sensations such as muscle length, muscle tension, and joint angulation. The cell bodies of the primary sensory neurons, which transmit all these sensations to the central nervous system, are located within the dorsal root ganglia of the spinal nerves (and, for these same senses in the head, in the sensory ganglion of the trigeminal nerve). The axons of these sensory neurons enter the central nervous system by the dorsal roots of the spinal nerves (and the afferent root of the trigeminal nerve). The axons branch upon entering the central nervous system. Some of these branches synapse onto interneurons within the adjacent gray matter; a subset of these interneurons in turn project onto motor neurons in the ventral gray matter of the same or neighboring segments. The axons of motor neurons leave the central nervous system and travel as a component of peripheral nerves to reach and synapse upon skeletal muscle in the periphery. The complete route just described—from peripheral somatic receptors to somatic afferent pathway to interneuron to motor neuron to peripheral motor neuron axon to muscle—forms the neural circuit that provides the anatomic basis for local reflex responses. (For only the myotatic reflex, also termed the tendon jerk, the interneuron is not present in this circuit; see Fig.
8.5.)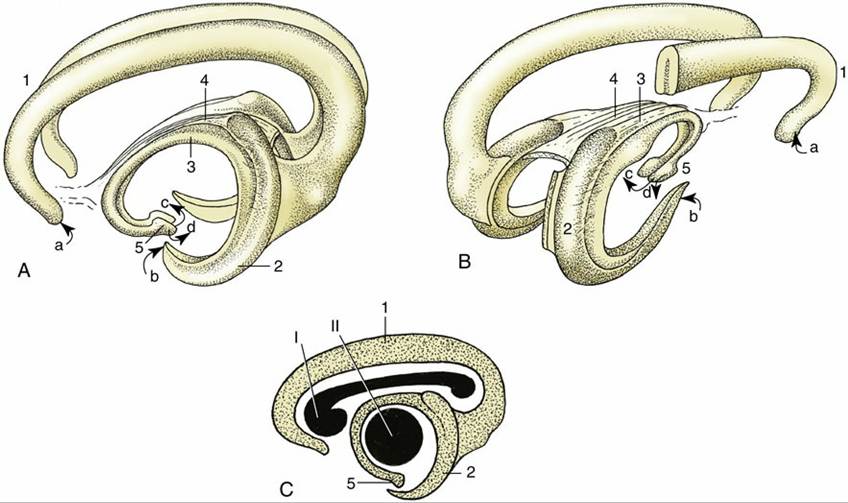
FIG. 8.38 Three-dimensional representations of the archipallium. (A) Left lateral view and (B) right caudolateral view. (C) The positions of the corpus callosum (I) and the thalamus (II) are shown in lateral projection. 1, Supracallosal and cingulate gyri; 2, hippocampus; 3, fornix; 4, commissure of fornix; 5, hypothalamus with mammillary body. a, Input from the medial olfactory tract; b, input from the piriform
lobe; c, output to the mammillothalamic tract; d, output to the brainstem.
Other branches of the primary sensory axons connect directly, or through interneurons, with higher centers in the brain or spinal cord, thus providing pathways that initiate more complex integrated responses. These ascending pathways, described in more detail later, form some of the white matter tracts of the spinal cord. Once in the brain, these pathways ultimately reach the somatosensory area of the cerebral cortex, providing the mechanism for conscious perception of these various somatic sensations. Additionally, as these pathways ascend to and through the brain, they produce collateral branches at different levels to connect to numerous brain regions.
The Lemniscal System
There are two main ascending pathways that reach the somatosensory region of the cerebral cortex. The first, termed here the lemniscal system though other names are used, transmits information that provides a high degree of spatial discrimination of touch, an accurate assessment of the intensity of pressure, repetitive vibratory sensation, and muscle and joint proprioception. Spinal interneurons are not involved in this pathway. Instead, some of the axonal branches of the primary sensory neurons, after entering the spinal cord, pass at once to the dorsal funiculus of the cord, that component of the spinal white matter located dorsally (see Fig. 8.7). These primary sensory axons adopt a very orderly arrangement within the dorsal funiculus (see Fig.
8.18); those that enter through sacral nerves—that is, most caudally—occupy the most medial positions, whereas those that enter at more cranial levels assume progressively more lateral positions. A glial septum that appears within the dorsal funiculus at midthoracic level divides it into two parts: the medial division, which constitutes the gracile fasciculus, contains axons from the hindlimb and caudal trunk; the lateral division, the cuneate fasciculus, contains axons from the forelimb, the cranial part of the trunk, and the neck. Both tracts end within corresponding nuclei, gracile and cuneate nuclei, within the dorsal part of the medulla oblongata, where they raise slight surface elevations, the gracile and cuneate tubercles (Fig. 8.22/13 and 14). The axons of the second-stage neurons—that is, neurons located within the gracile and cuneate nuclei—leave the ventral aspects of the nuclei and at once decussate, or cross, to the opposite side and turn rostrally as the large axon tract known as the medial lemniscus. The medial lemniscus runs cranially within the ventral part of the medulla, dorsal to the pyramid and close to the midline, to reach a specific part of the caudoventral nuclear complex of the thalamus (MCV) (Fig. 8.41). After synapses within the thalamus, axons of third-stage neurons travel within the white matter projection termed the thalamic radiation to the somatosensory area of the cerebral cortex (neopallium) (see Figs. 8.20B/2,3 and 8.23B/2,3). This is a cortical area directly caudal to the cruciate sulcus. In addition to information transmitted via spinal somatic sensory pathways, the lemniscal pathway receives somatic sensation from the head and neck at the level of the brainstem, where the medial lemniscus is joined by axons transmitting somatic sensory information from the lateral cervical nucleus, the nucleus of the descending tract of the trigeminal nerve, and the rostral (principal) sensory nucleus of the trigeminal nerve after a decussation within the metencephalon (Figs. 8.41 and 8.42).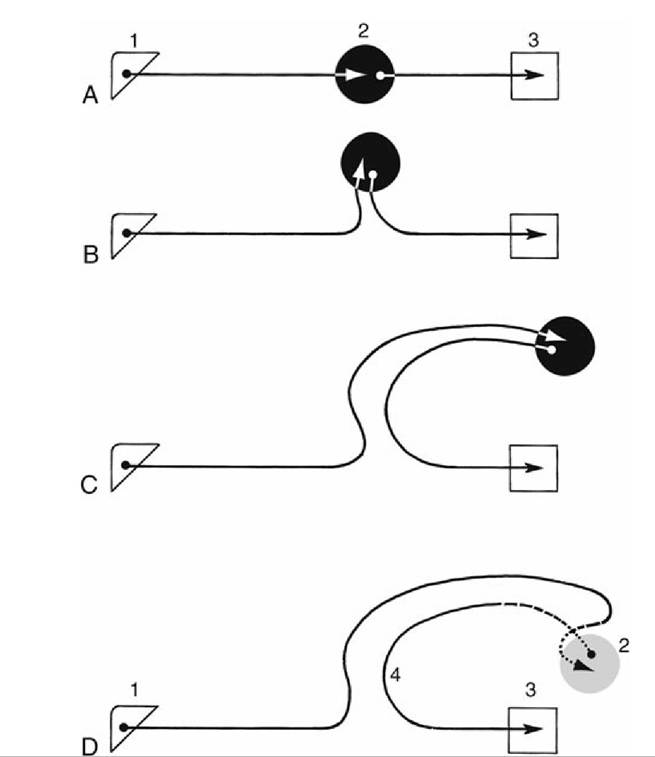
FIG. 8.39 (A) Diagram illustrating conjectured course of fibers running to and from the hippocampus. Because of differential growth of various parts of the brain, the hippocampus extends first (B) dorsally, then (C) caudally, and finally (D) laterally. 1, Olfactory bulb; 2, hippocampus; 3, hypothalamus; 4, fornix.
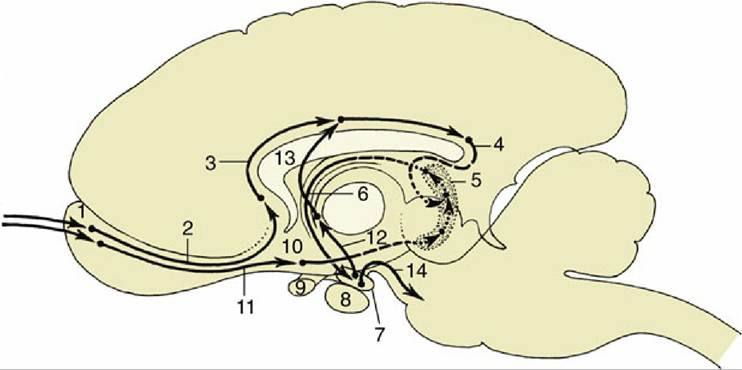
FIG. 8.40 A simplified conjectured diagram of the relay scheme of the limbic system. The solid arrows indicate the relay of information from the olfactory bulbs through the structures of the limbic system. The fiber tracts indicated by dotted lines are bent laterally out of the plane of the drawing. 1, Olfactory bulb; 2, medial olfactory tract; 3, cingulum (in gyri supracallosus and cinguli); 4, gyrus dentatus; 5, hippocampus; 6, fornix; 7, mammillary body; 8, hypophysis; 9, optic chiasm; 10, piriform lobe; 11, lateral olfactory tract;
12, mammillothalamic tract; 13, projection fibers entering the cingulum; 14, projection fibers to reticular formation.
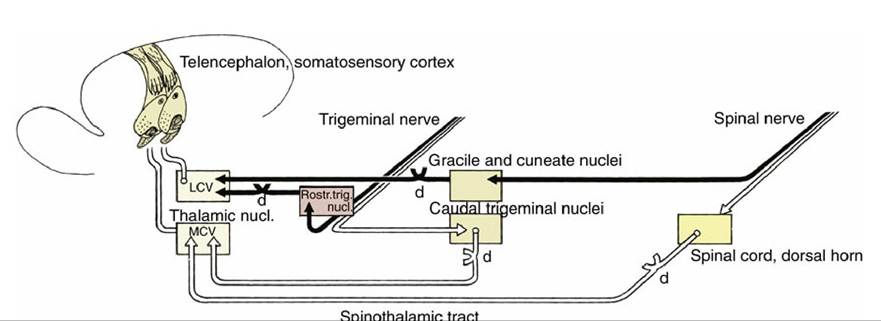
FIG. 8.41 The lemniscal (black) and extralemniscal (white) projections from the trunk and head to the telencephalon. d, Decussation; LCV, lateral part; MCV, medial part of the Caudoventral thalamic nucleus.
The somatotopic organization of the lemniscal pathway is preserved throughout its length, including the thalamic nucleus and the cortex. The somatotopic representation of the body within the cortex is of contralateral parts of the body and reflects the density of the sensory innervation of each body region rather than its absolute size. There is also some segregation within the cortex by sensory modality.
The Extralemniscal System
The extralemniscal system transmits a second subset of somatic afferent modalities, characterized by slower propagation and less precise localization of the originating stimuli.
The information conveyed is composed of cruder varieties of touch and pressure sensation, temperature sensation, and, most important, pain sensation. Within the spinal cord, branches of the primary sensory axons of this system synapse on interneurons located in the dorsal gray matter within a segment or two of axonal entry into the spinal cord. The information is processed via several spinal interneurons before leaving the dorsal horn (see Fig. 8.7). The axons of these second-stage neurons then pass into the white matter of the cord and ascend to higher brain centers. The projection of pain signals in particular from the spinal cord to the brain occurs via multiple ascending systems, which can be divided into medial and lateral groups by their projections.The tracts of the medial group tend to project and synapse upon regions throughout the brainstem up to the level of the diencephalon. This group is made up of the spinothalamic tract (see Fig. 8.42) that projects to the medial and intralaminar thalamic nuclei; the spinoreticular tract, composed of axon bundles located bilaterally within the ventral and ventrolateral regions of the spinal white matter that synapse within the reticular formation of the brainstem as far rostrally as the thalamus; and a loosely organized group of ascending propriospinal axons that originate and end repeatedly in the spinal gray matter, forming a multisynaptic relay system for information to ascend to the brain. These three main components of the medial group show little variation among vertebrates, in contrast to the lateral group described next.
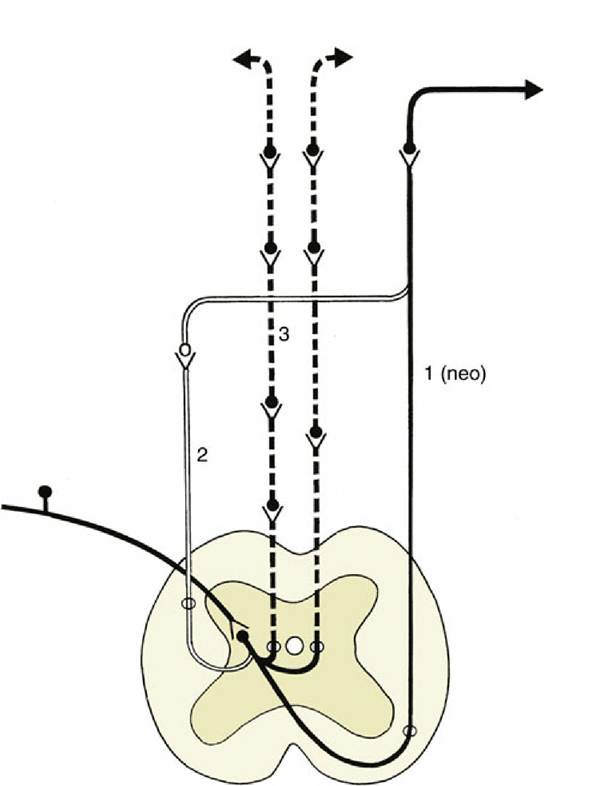
FIG. 8.42 A simplified scheme of the extralemniscal projections ascending from the spinal cord to the telencephalon. The solid black and white lines represent the projections within the lateral system; the interrupted black lines represent the bilateral and multisynaptic projections within the medial system.
The (paleo) spinothalamic tract is not represented in this scheme. 1, Spinothalamic tract; 2, spinocervicothalamic tract; 3, spinoreticulothalamic tract.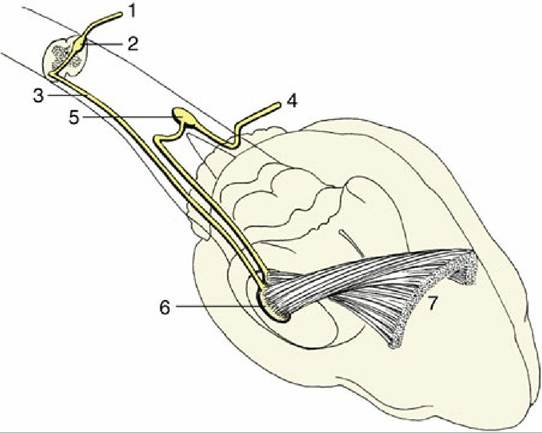
FIG. 8.43 Three-dimensional representation of the extralemniscal projection in the dog. 1, Spinal nerve;
2, dorsal horn of spinal cord; 3, spinothalamic tract; 4, trigeminal nerve; 5, nucleus of the spinal tract of the
trigeminal nerve; 6, medial part of the Caudoventral thalamic nucleus; 7, somatosensory cortex.
The lateral group is made up of tracts projecting onto the medial caudoventral nuclear complex of the thalamus, similar to the lemniscal pathways. The tracts making up the lateral group include the (neo)spinothalamic tract, the spinocervicothalamic system, and the second-order dorsal column pathway (Figs. 8.42 and 8.43).
The (neo)spinothalamic tract constitutes the classic pain tract of primates, including humans. It is entirely crossed and ascends within the spinal cord white matter on the ventrolateral aspect of the spinal cord and then through the brainstem toward the MCV.
The spinocervicothalamic system is well developed in subprimate mammals, particularly carnivores. Second-order axons arising from spinal interneurons ascend ipsilaterally as the spinocervical tract, located in the dorsolateral region of the spinal white matter, and synapse in the lateral cervical nucleus, located at the junction of the spinal cord and brainstem. The third-order axons that arise from this nucleus cross the midline and follow the medial lemniscus to end in the MCV, where they overlap the projection site of the (neo)spinothalamic tract.
The third system has been found in cats. It is composed of second-order axons from spinal interneurons that, surprisingly, ascend as a component of the spinal dorsal columns alongside the axons of primary afferent neurons of the lemniscal system. The postsynaptic, pain-conveying axons of this third system synapse in ipsilateral dorsal column nuclei. From neurons in the dorsal column nuclei, third-order axons cross the midline and also run to the MCV.
For sensory information from the head and neck traveling in the extralemniscal system, second- order axons arise from the caudal part of the descending trigeminal nucleus. These axons either join the lateral system and ascend to synapse in the MCV or join the medial system and ascend to the reticular formation of the thalamus. The third-order axons arising in the thalamus project to an area of the somatosensory cortex rostral to the area allocated to the lemniscal system.
Models have been proposed to explain the respective roles of the lateral and medial painsignaling systems in the generation of pain sensation and behavior. It has been proposed that the lateral and medial systems contribute differentially to the psychological dimensions of pain experience: one suggestion is that the lateral system conveys information regarding the sensory- discriminative dimensions of pain, whereas the medial is mainly involved in the motivational- affective dimension via the reticular formation, medial thalamus, and limbic system. Another model suggests that the lateral system is tuned preferentially to the sudden onset of noxious stimuli and thus may be related to the threat modality of pain. In contrast, the medial system is tuned to persistent components of pain and is thus better suited to mediate signals relating to existing tissue damage.
Subconscious Proprioceptive Pathways
The pathways transmitting information about the position and movement of the limbs — that is, proprioceptive information—to the somatosensory cortex have already been described as a component of the lemniscal system. An entirely different set of pathways transmits the same proprioceptive information to the brain but only as far as the cerebellum. These pathways are considered subconscious proprioceptive pathways as they do not terminate in the cerebral cortex. The pathways commence in the usual way with primary sensory axons arising from muscle and joint receptors that terminate on interneurons in spinal gray matter within the initial and adjacent spinal segments. The axons of the second-stage neurons travel cranially as the dorsal and ventral spinocerebellar tracts constituting the lateralmost regions of spinal white matter (Fig. 8.18/5 and 6). The dorsal tract takes a direct ipsilateral pathway that enters the cerebellum through the caudal cerebellar peduncle; the information it conveys is obtained from stimulation of muscle spindles. In contrast, the ventral spinocerebellar tract is concerned mainly with transmitting information arising from tendon receptors. The axons of the ventral tract decussate within the cord close to their origins; they then ascend to midbrain level before they turn caudally to enter the cerebellum through the rostral cerebellar peduncle. A second decussation within the cerebellar medulla restores the axons of the ventral tract to the side of the origin of the stimulus before they terminate within the cerebellar cortex. These two tracts transmit information only from the trunk and hindlimbs; the equivalent representation of the forelimb follows a different pathway that is not described here.
A further diffuse ascending pathway is provided within the reticular formation, the subject of the following section. It provides a means for integrating information conveyed by the pathways previously described with information from other afferent systems, somatic and visceral, general and special.
The Reticular Formation
The reticular formation extends throughout the brainstem as a diffuse arrangement of neurons interspersed with axon tracts. In the evolutionary sense, it is an old system. Despite the lack of obvious organization, closer analysis permits the recognition of numerous nuclei of varying size and architecture; some are sufficiently distinctive for their homologues to be recognizable in different species.
The reticular formation is connected to all projection systems within the central nervous system, whether afferent or efferent, and has reciprocal connections with the major integration centers within the brain. Thus, among its many ascending, descending, and transverse connections, there are such tracts as reticulocerebellar and cerebelloreticular and reticulothalamocortical and corticoreticular. The inescapable inference is that the reticular formation plays an important role in modulating the activities of these integration centers.
The reticular formation occupies a large part of the brainstem; it forms the main substance of the medulla, pons and midbrain, and when it reaches the thalamus, it contributes some of the nuclear groups of this complex structure. It also extends into the cervical part of the spinal cord.
The reticular formation may be divided into parts distinguished by morphology and location. The medial part, the periventricular gray, is located adjacent to the ventricular system of the brain. It has proved impossible to analyze in detail but appears to provide multisynaptic pathways composed of an indeterminable number of neurons with short and much branched processes. The second component exhibits a more obvious organization with more readily identifiable nuclei and ascending and descending tracts. For example, the reticular nuclei of the thalamus receive an input from lower parts of the reticular formation as the Spinoreticulothalamic tract and project diffusely on the entire neopallium. This tract is an important component of the reticular system and may be a route, complementary to the spinothalamic tract, for somatic sensory information to reach the cortex. It contains axons that project for long distances and conduct more rapidly than those found in the spinothalamic tract.
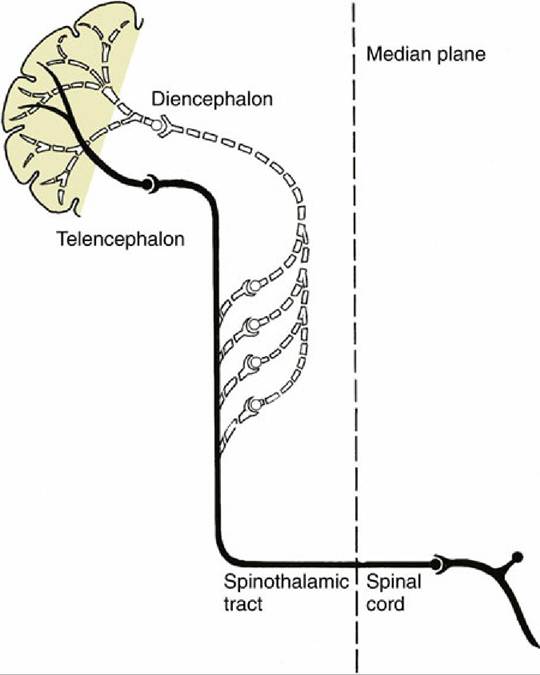
FIG. 8.44 A multisynaptic ascending tract (white dotted line) to the telencephalon via the reticular formation. The collateral tract in this example represents the extralemniscal projection (solid black line).
One extensive reticular ascending pathway that ultimately projects beyond the thalamus to the cortex is known as the ascending reticular activating system (ARAS). The ARAS receives inputs through collateral branches from all sensory systems, whether somatic or visceral (Fig. 8.44). Its activation arouses the animal, making it more conscious of its circumstances and surroundings; diminution of ARAS activity induces lethargy or sleep. The ARAS has been regarded as an important component of the seat of consciousness, although most neurologists would assert that "there is no single place where consciousness dwells."
The reticular system also plays an essential role in motor control by means of descending pathways from the telencephalon that synapse on reticular nuclei. In turn, these nuclei project axons to synapse on lower motor neurons of the brainstem and cord.