The Internal Ear
The mechanical stimuli produced by sound and by the positional changes of the head are transformed into nerve impulses in the internal ear. This is a delicate mechanism, no larger than about 12 mm across in the dog, and is completely enclosed in the very hard petrous temporal bone for protection and proper functioning (see Fig.
9.24A). It is exposed to sound vibrations on the lateral surface, and the electrical impulses into which these vibrations are converted leave the internal ear in nerve fibres of the vestibulocochlear nerve, which passes through the internal acoustic meatus on the medial surface.The internal ear consists of a closed system of tiny membranous ducts and cavities known, because of its complexity, as the membranous labyrinth (Fig. 9.28A). It contains endolymph whose movement inside the system stimulates sensory cells in the membranous wall. Two enlargements in the center of the membranous labyrinth are known as the utriculus and sacculus. From the utriculus arise three semicircular ducts, and the sacculus is attached to the spiral cochlear duct. Although these structures are all closely related to each other developmentally and anatomically, their functions are distinct. The utriculus, sacculus, and semicircular ducts are concerned with detection of head position and movement, whereas the cochlear duct is concerned with hearing.
The semicircular ducts stand roughly at right angles to one another and are designated anterior, posterior, and lateral; one end of each duct is widened, or ampullated, close to the utriculus. The endolymph within the ducts is set in motion by movements of the head, applying force on receptive structures known as ampullary crests in each ampulla (Fig. 9.28/9 and 10). An important component of the ampullary crests are hair cells, so-called because of their cilia, which project into the ampulla.
Forces exerted by the motion of the endolymph cause the cilia to bend, stimulating the hair cells to release neurotransmitter onto closely associated sensory neurons of the vestibular portion of the vestibulocochlear nerve, which in turn send impulses to the central nervous system.Two further receptor areas called maculae (Fig. 9.28/6 and 7) are present in the walls of the utriculus and sacculus. They monitor the position of the head with respect to gravity. Like the ampullary crests, the maculae are bathed in endolymph and also contain hair cells. Instead of reacting to movement of the endolymph, however, the cilia of the macular hair cells are embedded in a gelatinous substance in which a layer of crystals (statoconia) is adhered. When the gelatinous layer of the maculae faces toward the ground, the cells are maximally stimulated by the gravitational pull on the statoconia. The maculae detect the position and linear movements of the head, whereas the ampullary crests detect the rotational movements of the head.
The sacculus gives origin to the endolymphatic duct, which ends blindly in the epidural space (Fig. 9.24/17). It is thought to function in the resorption of the endolymph secreted by the epithelial lining of the membranous labyrinth.
The membranous labyrinth is housed in a similar but slightly larger osseous labyrinth, a complex excavation in the temporal bone. The central chamber of the osseous labyrinth, the vestibule, houses the utriculus and the sacculus. The semicircular ducts lie within the osseous semicircular canals. The cochlear duct lies within the bony spiral canal of the cochlea, which is an excavation very similar to the inside of a snail's shell. The center of the cochlea is an osseous pyramid known as the modiolus (Fig. 9.29/2). Running around the modiolus is the spiral canal, the actual lumen of the cochlea, which ends blindly at the apex of the modiolus. Projecting into the spiral canal from the modiolus is an osseous shelf, the spiral lamina (Fig.
9.29/5), which terminates in the blind end of the spiral canal of the cochlea. The spiral lamina itself is hollow, forming the spiral canal of the modiolus.Because the osseous labyrinth is slightly larger than the membranous labyrinth it encloses, there is a minute space between the two that contains perilymph. The perilymphatic space is only of significance in the area associated with the cochlea, where it forms two chambers, the scala tympani and the scala vestibuli, described below.
The bony spiral canal of the cochlea is divided into three channels (Fig. 9.29/A6-8), by a split longitudinal membrane, all running around the modiolus to the apex of the cochlea. The membrane arises centrally from the spiral lamina and, after splitting, attaches to the outside wall of the spiral canal. The uppermost channel is the scala vestibuli (Fig. 9.29/6), the middle one is the cochlear duct (Fig. 9.29/7), and the lowest is the scala tympani (Fig. 9.29/8). The two scalae communicate at the apex of the cochlea around the blind end of the cochlear duct. At the base of the cochlea, the scala vestibuli communicates with the perilymphatic space in the vestibule, and the scala tympani ends at the secondary tympanic membrane of the cochlear window (described previously; see Fig. 9.24).
An enlarged transverse section of the spiral canal of the cochlea shows the composition of the split membrane, particularly the part that forms the walls of the triangular cochlear duct (Fig. 9.29/7). The simplest of these walls separates the cochlear duct from the scala vestibuli; it consists of a single layer of cells and is known as the spiral membrane (Fig. 9.29A/12). The wall of the cochlear duct facing the scala tympani is complex by virtue of an arrangement of multiple rows of hair cells and other cells found in it, termed the spiral organ (Fig. 9.29A/13). Its connective tissue base is the basilar lamina, the characteristics of which play an important role in the transduction of different sound frequencies.
The functioning of the cochlea is described here briefly. In the middle ear, the mechanical movements of the base of the stapes vibrate the vestibular window, as described previously. Movement of the vestibular window in turn compresses the perilymph in the closed system of perilymphatic spaces in the inner ear. Because fluids are incompressible, pressure exerted on the fluid in the scala tympani results in a pressure wave traveling up to the apex of the bony cochlea. As the pressure wave passes, the force serves to move the pliable basilar lamina of the cochlear duct. This movement bends the cilia of the hair cells in the basal lamina, causing release of neurotransmitter onto closely associated sensory neurons associated with the cochlear portion of the vestibulocochlear nerve. The details of how the movements of the basilar membrane act on the hair cells is complex and beyond the scope of this book. Suffice it to say that the width and compliance of the basilar lamina results in transduction of lower frequency waves by the spiral organ located near the apex of the cochlea and that of higher frequency sounds by more proximal regions of the basilar membrane and spiral organ.
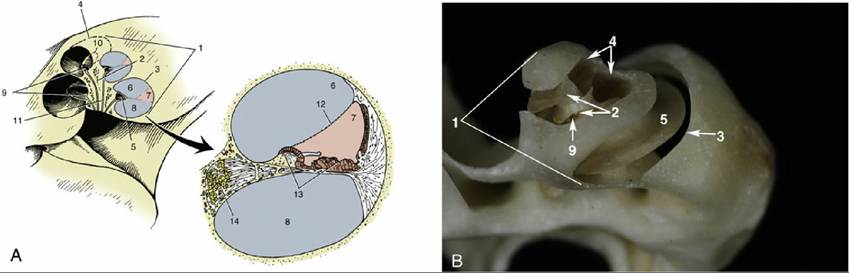
FIG. 9.29 Cochlea and enlarged cochlear duct. Schematic (A) and of the dog (B). 1, Cochlea; 2, modiolus; 3 and 4, spiral canal of cochlea; 5, osseous spiral lamina; 6, scala vestibuli; 7, cochlear duct; 8, scala tympani; 9 and 10, spiral canal of modiolus; 11, longitudinal canals; 12, spiral membrane; 13, spiral organ; 14, spiral ganglion.
The impulses generated in cochlear fibers travel toward the modiolus to ganglion cells housed in the spiral canal. The aggregate of these cells forms the spiral ganglion (Fig. 9.29A/14), which also winds around the modiolus. From the spiral ganglion, the impulses travel along nerve fibers within canals to the base of the modiolus, where the fibers join to form the cochlear part of the vestibulocochlear nerve.
The vestibulocochlear nerve (cranial nerve VIII) is composed of vestibular and cochlear parts as it enters the internal acoustic meatus. As previously mentioned, the branches of the vestibular portion arise from the maculae in the utriculus and sacculus and the ampullae in the semicircular canals, conveying impulses concerned with balance; the cochlear portion arises from the base of the cochlea to mediate the impulses concerned with hearing.
The anatomy of the internal and middle ear is complicated by the passage of the facial nerve through this area (Fig. 9.24/10). The facial nerve enters the internal acoustic meatus together with the vestibulocochlear nerve and, within an osseous facial canal, traverses the temporal bone to emerge at the stylomastoid foramen. The facial canal makes a sharp kneelike bend within the temporal bone, and at this point the nerve is enlarged by the geniculate ganglion. From this ganglion arises the major petrosal nerve, which regulates secretion of the lacrimal and nasal glands. The chorda tympani, regulating the sublingual and mandibular glands but also relaying taste from the rostral two thirds of the tongue, leaves the facial nerve a little more distally. The chorda tympani is so named because, for a short segment of its course, it lies on the upper part of the tympanic membrane (Fig. 9.26/5). Both major petrosal and chorda tympani nerves leave the temporal bone through foramina on the rostroventral aspect of the bone. The facial nerve also supplies the stapedius muscle. (The tensor tympani is activated through the mandibular division of the trigeminal nerve [V3].)