THE LUNGS
The right and left lungs (pulmones, pl.) are each invag- inated into the corresponding pleural sac and are free, except at the roots where they are attached to the mediastinum. They have no fixed size or shape since they comply with respiratory changes in the dimensions of the thorax.
The lungs are normally kept expanded by the air pressure within the respiratory tree, and being elastic, they recoil and collapse as soon as air is admitted into the pleural cavities by trauma, surgery, or dissection. They have a soft, spongy texture, and the residual air they contain, even when collapsed, causes them to crepitate when squeezed and to float when placed in water. In contrast, the unexpanded lungs of the fetus or stillborn animal feel solid; they sink when immersed, and this provides the pathologist with an easy means of determining that the animal from which they came had not breathed. The color of healthy lungs varies in intensity with the blood content and therefore with the manner of death; it is a fresh pink in many slaughterhouse specimens but a much deeper red in lungs obtained from animals that were not bled. The frequently patchy coloration is produced by uneven distribution of blood, which is often the result of gravitation after death. The lungs of animals that spent their lives in heavily polluted atmospheres acquire a grayish tinge from deposition of soot or other inhaled particles.Anatomical descriptions are generally based on specimens hardened in situ before the thorax was opened; at death such lungs retain their size, which is intermediate between those adopted in full inspiration and full expiration (Figure 4-22). The two lungs are grossly alike and mirror each other in shape, although the right one is always larger; this asymmetry, partly due to the skewed position of the heart, is most obvious in the lungs of cattle.
Each has some resemblance to the half of a cone, which makes it possible to recognize the following features: an apex presented toward the thoracic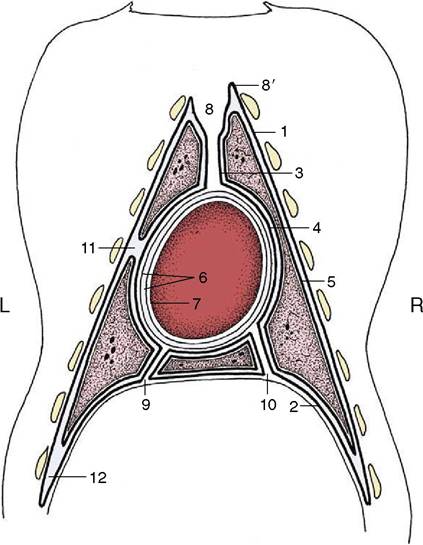
Figure 4-21 The distribution of the pleura and pericardium, schematic. The heavy lines indicate the pleura. 1-4, Parietal pleura, later subdivided; 1, costal pleura; 2, diaphragmatic pleura; 3, mediastinal pleura; 4, pericardial pleura; 5, visceral (pulmonary) pleura; 6, parietal pericardium; its outer fibrous layer tightly adheres to its inner serous layer; 7, visceral pericardium, adherent to heart (epicardium); 8, cranial mediastinum; 8', cupula pleurae; 9, caudal mediastinum; 10, plica venae cavae; 11, left cardiac notch; 12, costodiaphragmatic recess.
inlet; a wide, concave base related to the face of the diaphragm; a convex costal surface fitted against the lateral chest wall; an irregular medial surface modeled on the contents of the mediastinum; a thick dorsal border occupying the gutter between the vertebrae and ribs; and a thin border that comprises a ventral part bordering the costomediastinal recess and a basal (cau- doventral) part bordering the costodiaphragmatic recess (Figures 4-20 and 4-22). The ventral part is indented over the heart (cardiac notch; incisura cardiaca).
Certain features of the mediastinal surface and base require further attention. The many indentations carried by the mediastinal surface include the large and deep cardiac impression, which is created by the heart and naturally larger on the left lung because the heart itself is biased to this side. The impression extends to the ventral border, which is deeply notched at this level in most species and which, in turn, allows the heart (or more accurately, the pericardium) direct contact with
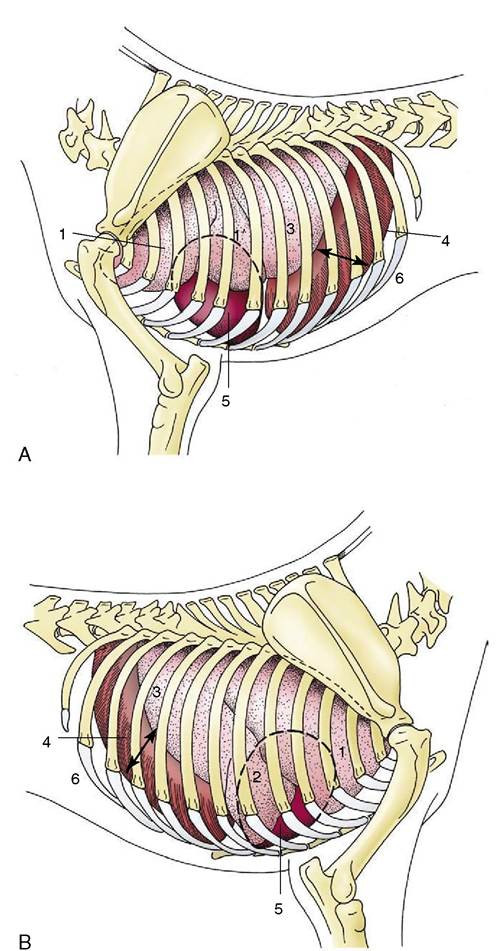
Figure 4-22 Semischematic drawings of the thoracic organs of the dog on the left (A) and right (B) sides.
The outline of the heart is indicated by a broken line. 1, Cranial lobe; 1, caudal part of left cranial lobe; 2, middle lobe; 3, caudal lobe; 4, diaphragm; 5, heart; 6, costodiaphragmatic recess (arrow).the thoracic wall (see Figure 4-22). The root of the lung, situated dorsal to the cardiac impression, is formed by the bunching together of the chief bronchus and the pulmonary artery, veins, lymphatics, and nerves within a covering of pleura provided by the reflection of the mediastinal pleura onto the lung. The reflection extends caudal to the root in a tapering fashion that leaves bare an area of lung that is directly joined by mediastinal connective tissue to the corresponding part of its partner. In some species, including the dog and the cat, the empty part of the reflection, which is known as the pulmonary ligament, extends onto the base of the lung, which thus finds additional attachment to the diaphragm. In ruminants and pigs the bronchus that arises from the trachea before its bifurcation together with the associated vessels creates a smaller second root of the right lung (Figure 4-23 and Figure 4-17, B).
The base of the right lung reveals the small accessory lobe, which is separated from the medial surface of the caudal lobe by a fissure that widens at its dorsal limit to accommodate the caudal vena cava in its passage between the caval foramen of the diaphragm and the right atrium. The accessory lobe sits, as it were, astride the vein.
In most species one or more fissures extend into the substance toward the root, dividing each lung into parts that are commonly equated with lobes. The lobes are properly defined by the ramification of the bronchial tree, and scope for confusion exists because many older texts employed the external demarcations for this purpose. According to the current practice, the left lung consists of cranial and caudal lobes and the right one of cranial, middle, caudal, and accessory lobes; however, the cranial lobe is commonly subdivided by an external fissure, whereas the right lung of the horse lacks a middle lobe.
The fissures are much deeper in the lungs of the dog and the cat than in those of other species, but it is difficult to find convincing functional significance in such differences. The deeper fissures may allow the parts to slip over each other more easily and facilitate the adaptation of the lungs to the pronounced changes in thoracic form that occur in animals that employ a bounding gallop.The bulk of the lung substance is provided by the bronchi, pulmonary vessels, and peribronchial and perivascular connective tissue. The right and left chief bronchi arise at the tracheal bifurcation above the heart, and after entering the lung at its root, each detaches a bronchus to the cranial lobe before continuing caudally (see Figures 4-17 and 4-24). The two generations of subdivisions that follow next have a fairly consistent pattern of origin, but subsequent ramifications are less predictable. The number of bronchial generations before the smaller bronchi are succeeded by bronchioles varies among species and also among parts of the one lung. In mice and other small animals only four or five generations of bronchi are present, whereas more than a dozen may be necessary in large animals. The consistency in the pattern of the first branchings allows the recognition of the so-called bronchopulmonary segments, specific portions of the lung supplied by identifiable bronchi and partly defined by connective tissue septa that extend from the peribronchial and perivascular tissue (and are responsible for the surface marbling where they impinge on the visceral pleura). Although bronchopulmonary segmentation has been studied in domestic species, it has yet to find important application; it is not yet common veterinary practice to resect portions of diseased lungs. It is the elasticity of the connective tissue stroma that allows the lungs to expand on inspiration and collapse on subsequent expiration. Loss of this elasticity, which occurs naturally with aging (but also in certain pathological conditions), reduces respiratory efficiency.
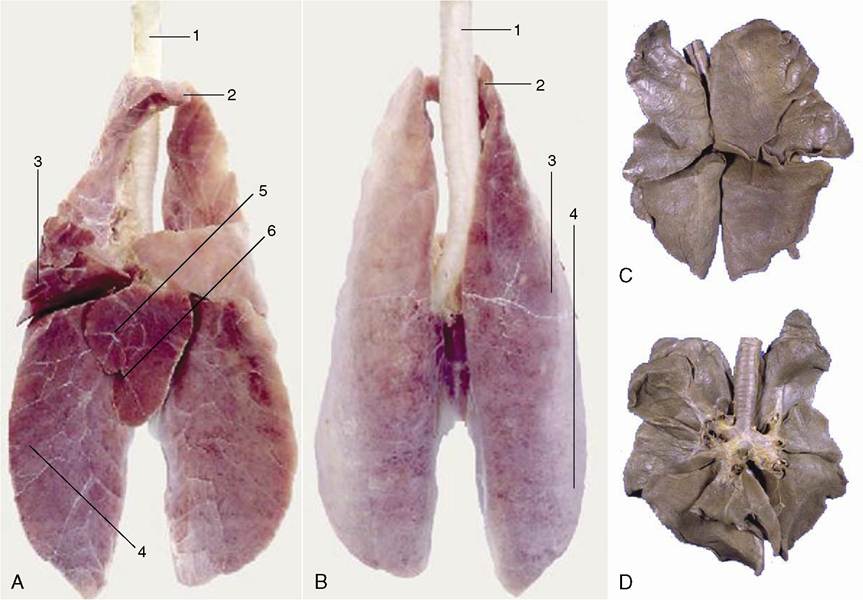
Figure 4-23 Ventral (A) and dorsal (B) surface of the lung of a pig. C, dorsal and D, ventral surface of the lung of a dog. Notice the deep indentations between the lobes of the canine lung. 1, Trachea; 2, cranial lobe; 3, middle lobe; 4, caudal lobe; 5, accessory lobe; 6, opening for caudal vena cava.
The structure of the major bronchi resembles that of the trachea, but with each successive division the supporting cartilages become smaller and more irregular, while the muscle expands to enclose the lumen on all sides. The lumen is lined by a pseudostratified epithelium comprising tall ciliated columnar cells interspersed with goblet and serous-secreting cells and with stem cells that proliferate to repair depletions of the other types. Larger glands are included within the submucosa of the major bronchi. The transition from bronchus to bronchiole is defined by the disappearance of the last cartilage plate and by the submucosal glands. Bronchioles are narrow—less than 1 mm in diameter—and also pass through several generations. The last of these is characterized by the loss of goblet cells and their replacement by the exocrinocytes (Clara cells) thought to secrete a component of lung surfactant. The terminal bronchioles present scattered alveolar outpouchings of their walls (and are continued by alveolar ducts), thence alveolar sacs, and ultimately by the saclike alveoli, the spaces where gaseous exchange takes place through a flattened epithelium closely related to the pulmonary capillaries. Patency of the finer passages, which are unsupported by cartilage, is ensured by elastic fibers that anchor them to the pulmonary stroma. At the first breath, the alveoli fill with air and dilate, although for a time they remain significantly smaller than those of the adult (Figure 4-25).
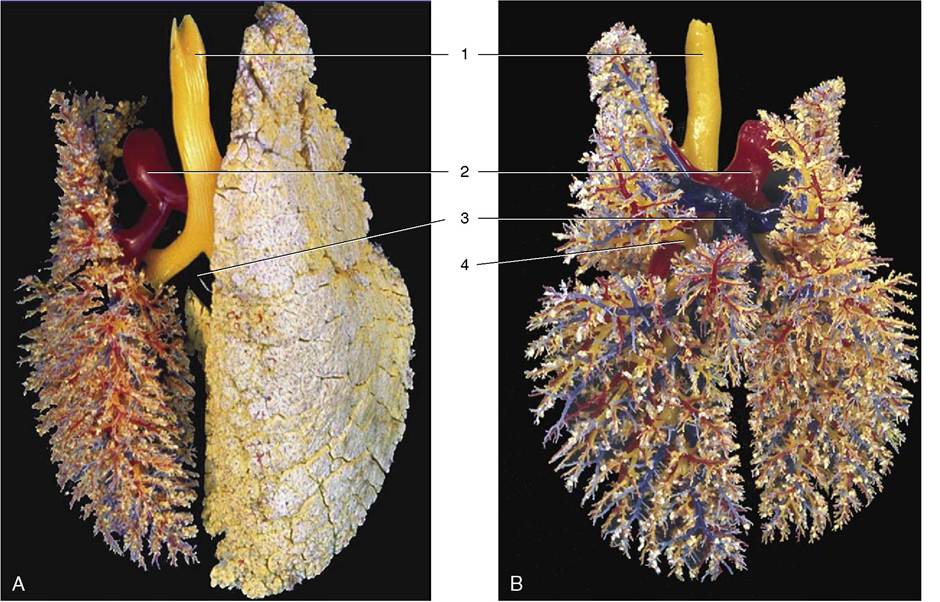
Figure 4-24 A, B, Dorsal view of the bronchial tree (yellow) and accompanying blood vessels of the pig (corrosion cast).
1, Trachea; 2, pulmonary trunk; 3, pulmonary veins; 4, tracheal bronchus.The identification of the lungs of individual species is most conveniently based on the degrees of lobation and lobulation. The lungs of horses show almost no lobation and very inconspicuous lobulation externally (Figure 4-26), those of ruminants (Figure 4-27) and pigs are conspicuously lobated and lobulated (though not uniformly in sheep and goats), and those of carnivores are very deeply fissured into lobes but show little external evidence of lobulation (see Figure 4-23).
The pulmonary arteries generally follow the bronchi (see Figure 4-24), while the pulmonary veins sometimes run separately, alternating in position with the broncho- arterial associations. The pattern varies not only with the species but also with location in the one lung. These
differences may find clinical significance if lung surgery becomes more common. Then it will be important to know the vascular arrangements and to be aware that both interarterial and intervenous anastomoses are to be found crossing the connective tissue partitions. A set of bronchial arteries arises from the aorta to supply the bronchi and associated connective tissue wholly independently of the pulmonary arteries (Figure 4-28). A corresponding set of bronchial veins may return this blood to the right atrium via the azygous vein, but often the bronchial flow is entirely returned to the left atrium. Arteriovenous anastomoses appear to be absent, and this makes the lung an effective filter for preventing the further spread of emboli and tumor cells. This accounts for the frequent occurrence of abscesses and tumor metastases in lung tissue, secondary to disease of other organs.
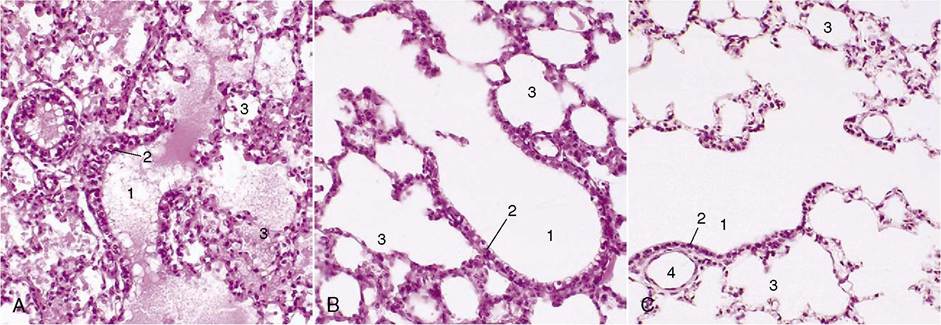
Figure 4-25 A, Lung of pig fetus (140?); note presence of fluid in bronchioles and alveoli. B, Lung of 1-day old piglet (140?).
C, Lung of an adult pig (140?). 1, Terminal bronchioles; 2, bronchiolar exocrinocyte (Clara) cells; 3, alveolar sac; 4, bronchiole.
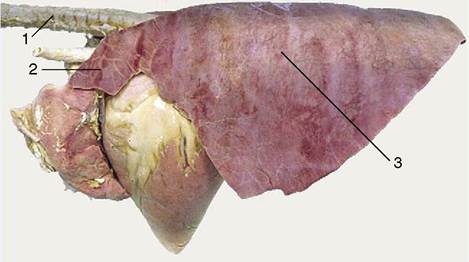
Figure 4-26 Left lateral view of the equine lungs. Note the poor lobation and lobulation. 1, Trachea; 2, cranial lobe; 3, caudal lobe.
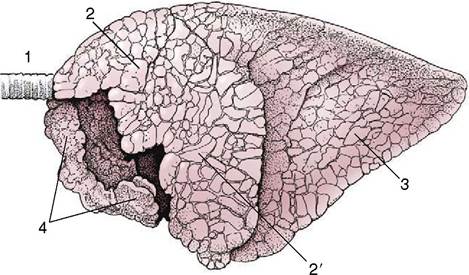
Figure 4-27 Left lateral view of the bovine lungs. Note the definite lobation and lobulation. 1, Trachea; 2, 2', cranial and caudal parts of left cranial lobe; 3, caudal lobe; 4, right cranial lobe.
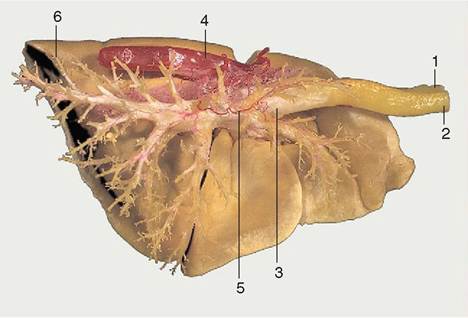
Figure 4-28 Corrosion specimen of the lungs and part of the aorta of a dog. On the right side the resin in the bronchioli and smaller bronchi has been removed to expose the main tracheobronchial tree. 1, Esophagus; 2, trachea; 3, tracheal bifurcation; 4, aorta; 5, bronchial artery; 6, caudal lobe of left lung.
Lymph drains to the tracheobronchial and mediastinal lymph nodes, directly or after initial passage through small pulmonary nodes set on the bronchial tree within the lung substance; the details are complicated, vary among species, and receive later notice when of pathological relevance.
The nerves to the lungs are delivered through a pulmonary plexus within the mediastinum to which both sympathetic and parasympathetic (vagal) fibers contribute. The efferent fibers pass to the bronchial glands and musculature and to the blood vessels. Afferent fibers come from the bronchial mucosa (cough reflex), from vessels, and from stretch receptors. Vagal section has been found to relieve pain in inoperable bronchial carcinoma of human patients.
The features of the lungs of greatest clinical significance are their projection on the surface of the body and their radiographic appearance. The projections vary among species and are described later; meanwhile, it may be stressed that they obviously vary with the phase of respiration. Moreover, the areas over which auscultation and percussion can usefully be employed are more limited than might initially be supposed; this is partly because intervention of the upper part of the forelimb denies access to part of the lung field and partly because the lower border of the lung is too thin to provide much useful information.
Because radiography of the lungs is done mainly in small animals (dogs and cats), the relevant observations on their appearance on radiographs and figures will be found in Chapter 13.
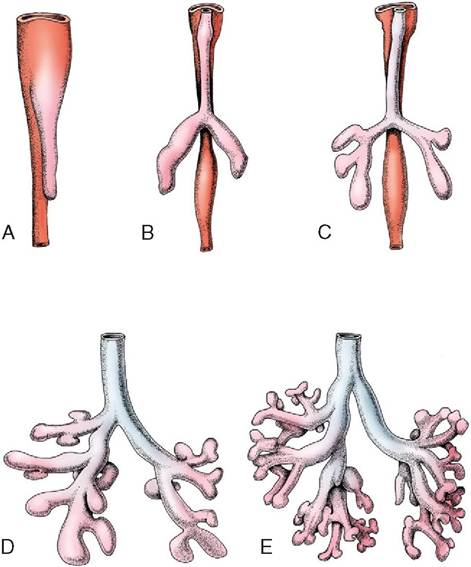
Figure 4-29 Five stages in the development of the trachea and lungs (ventral view). A, Caudal growth of the tracheobronchial tube. B, Its division into two lung buds. C, Further division into three bronchi on the right and two on the left. D, E, Further development of the bronchial tree.