The Motor Cortices of the Frontal Lobe, the Highest Level of the Motor Control Hierarchy, Consist of Three Different Functional Regions
The motor cortices of the frontal lobe, the origin of most of the corticospinal tract axons, are composed of the primary motor cortex, the supplementary motor cortex, and the premotor cortex (Figure 10-6; see also Figure IO-I).
Although these forebrain regions collectively represent the highest level of the motor control hierarchy, the areas appear to differ with respect to the complexity of motor functions controlled.In primates the primary motor cortex (MI) is located just rostral to the prominent central sulcus and therefore lies along the precentral gyrus (Figure 10-6, A). In many nonprimate mammalian species a central sulcus is not present, and MI appears to lie near the cruciate sulcus (Figure 10-6, B). Low- level electrical stimulation of a very small region of Ml is capable of activating a small number of functionally related muscles. Furthermore, an orderly relationship exists between the region of the body where the muscles are activated and the region of Ml stimulated. In this Somatotopic map of the body musculature in MI, muscles in the caudal part of the body (or the feet in bipeds) can be most easily activated from more
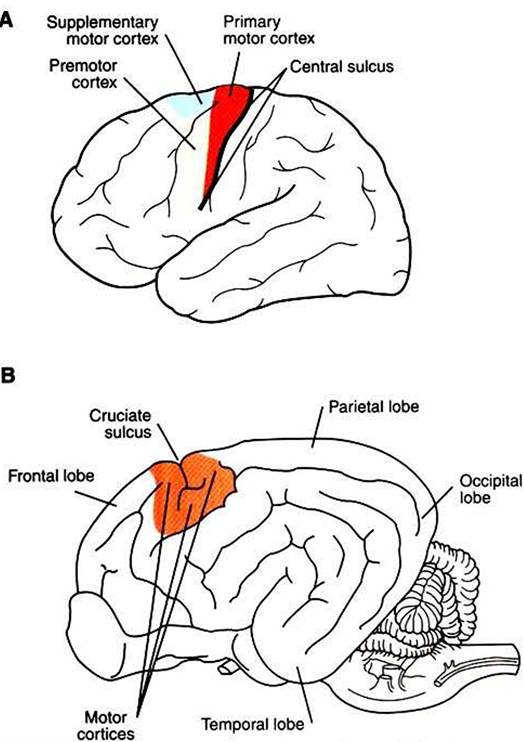
FIGURE 10-6 Motor cortices. A, Location of primary motor, supplementary motor, and premotor cortices in the human brain. BrVicinity of the motor cortices in the canine brain.
dorsomedial parts of MI, whereas muscles in the rostral part of the body (or head in bipeds) can be most easily activated from more ventrolateral parts of Ml, with a fairly orderly representation of the other regions of the body between those parts of MI. As shown in Figure 10-7, the musculature of different parts of the body is not equally represented in the Somatotopic map. Regions depicted as larger have a larger area of MI devoted to their voluntary muscular control, and thus the movements of that region will generally be that much more precise and fractionated.
In the somato- topic map of MI in humans, the hand and the mouth musculatures have a very large proportional representation, reflecting the respective importance of these areas in manipulating objects with the fingers and in articulating speech. The proportional representation of the musculature of the different body parts in Ml varies with phylogeny, but the somato- topic maps of the primates tend to be the most detailed, reflecting the most precise control over skilled, voluntary movements.The supplementary motor cortex and the premotor cortex are also located in the frontal lobe, just rostral to Ml, with sup-
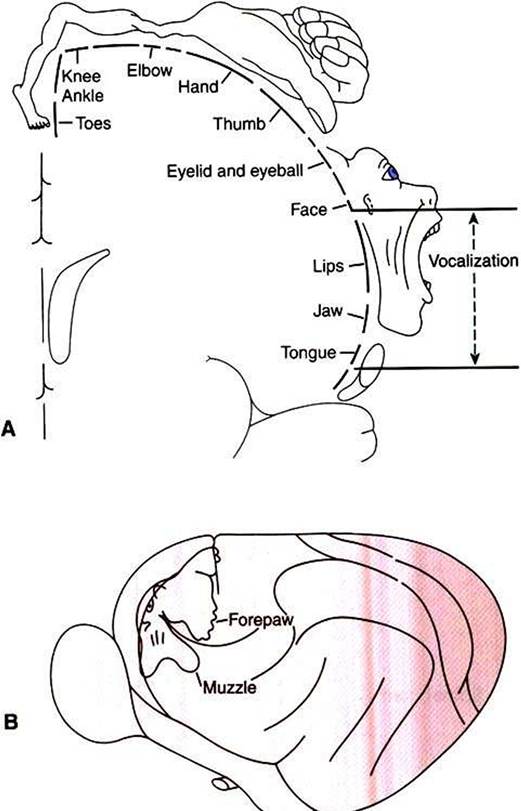
FIGURE 10-7 Somatotopic map of primary motor cortex (Ml) showing the origins of axons going to the different skeletal muscles of the body. Body parts represented as proportionally larger have a larger area of Ml devoted to their voluntary control, and the movement of that part will generally be that much more precise and fractionated. A, In the human, muscles controlling the hand/digits and the mouth are disproportionately represented because these muscles are needed for the critical and precise movements of grasping/manipulation and speech. B, Primary motor cortex of a cat. (A redrawn from PenfieIdW, RasmussenT: The cerebral cortex of man, NewYork, 1950, Macmillan; from Berne RM, Levy MN: Physiology, ed 2, St Louis, 1988, Mosby; B from Prosser CL: Comparative animal physiology, ed 3, NewYork, 1988, Wiley.)
plementary motor cortex positioned dorsomedial to premotor cortex (see Figure 10-6, Λ). Both areas also have a Somatotopic map of the body musculature, but it is less precise than in MLIn addition to corticospinal and Corticobulbar tract axons, both areas also collectively give rise to axonal projections to nuclei of origin of some descending brainstem motor pathways (see Figure 10-1). Most significantly, however, the supplementary motor and premotor cortices send axons to synapse within MI and thus may represent “supramotor” areas, with an even higher status in the motor control hierarchy than MI; these areas may instruct MI to organize its fairly discrete muscle actions into more elaborate movement patterns. This concept is supported by the fact that, for voluntary movement, neurons in these supramotor areas become active before those of MI.
Evidence indicates that the supplementary motor cortex is particularly important in the planning and organizing of complex sequences of the discrete movements normally carried out by MI. For example, supplementary motor cortex appears to be particularly active when an individual mentally rehearses a specific sequence of finger movements. Supplementary motor cortex also appears to be important for instructing the limbs (particularly the forelimbs) on the two sides of the body to work together, simultaneously, to accomplish a task. Premotor cortex appears to play an important role in preparatory orientation of the body for the execution of a particular motor task. An example in primates would be rotation of the shoulders and movement of the arms toward a target that is to be manipulated by the hands. Interestingly, both these areas receive integrated sensory input and Visuospatial information from the posterior parietal cortex, which most likely plays a role in the sequence-planning and preparatory orientation functions.
Using the analogy of playing the piano, we could view MI as being responsible for the simplest muscle activation necessary to press a single piano key, supplementary motor cortex as responsible for planning and organizing the sequence of such finger movements necessary to play a melody, and premotor cortex as responsible for orienting the arms and hands to the correct region of the keyboard to play the various sequences. Of course, the interaction among these areas in determining the appropriate corticospinal (and cortico- bulbar) tract activity necessary to produce the voluntary movement is certainly more complex than this, and the functional role of these areas in motor control, and how they work together, is still under investigation.
The severity of the deficits resulting from lesions of the corticospinal (pyramidal) system varies with the phylogenetic status of the animal. In primates, such as humans, in whom the pyramidal system is developed extensively, corticospinal tract lesions rostral to the pyramidal decussation cause a dense weakness of the contralateral side of the body.
Such one-sided weakness is called hemiparesis and is most extensive in the hand and facial muscles (symptoms common in ‘"stroke” in humans). These symptoms are understandable because a huge percentage of the corticospinal tract axons in primates cross the midline at the Spinomedullary border (the location of the pyramidal decussation), and the hand and face have the largest proportional representation in primary motor cortex (see Figures 10-4 and 10-7).In most veterinary species the corticospinal system is not as well developed as in humans, and supraspinal lesions of this system cause much less severe contralateral weakness and almost no alteration of gait. However, clinical examination can reveal more subtle deficits in voluntary movements of the contralateral limbs. An example is the proprioceptive positioning reaction, the ability of an animal to return its paw to a normal, pads-down posture after the clinician flexes it to make the dorsal surface touch the floor or tabletop. This response requires the animal’s conscious awareness that the paw is in the flexed position (conscious proprioception) and then requires that the animal be able to respond consciously by returning the paw to its normal posture. This latter motor response in turn is affected by the integrity of the upper motor neurons of the corticospinal tract. When these corticospinal tract neurons are damaged, the animal is slow to return its paw to a normal posture. In addition, toes tend to be dragged on the ground as the leg is drawn forward in normal gait. It should be noted that such deficits could also be produced by damaged Corticorubral axons originating in the motor cortices. Noting these conscious positioning deficits and other subtle gait changes is important in localizing lesions within the CNS.