THE NERVE IMPULSE AND ITS TRANSMISSION
1. What is the function of the Na+-K+ATPase pump in the axolemma?
2. What is the approximate value of the resting membrane potential?
3. What is the polarity of a resting neuron membrane?
4.
What accomplishes depolarization?5. What accomplishes repolarization?
6. Describe the sequence of events associated with neurotransmission in mammals.
7. What is an action potential?
8. What is threshold?
9. What is a refractory period?
0. What is meant by a nerve fiber being “fired”?
11. What is the “all-or-none” principle for nerve fibers?
2. How does neurotransmission differ in myelinated fibers?
3. What are two functions of saltatory conduction?
4. What kind of nerve fiber has the fastest impulse transmission?
L5. What is the purpose of a neuro transmitter?
6. What are the neurotransmitters associated with the autonomic nervous system and where are they located?
1.7. What is the nature of the central neurotransmitters?
8. Describe the final common pathway concept.
9. Differentiate between the different types of neuron circuits.
:0. What is the minimum number of neurons required for the transmission of a nerve impulse from the periphery by way of a spinal nerve to the cerebral cortex?
Communication among neurons and with the cells of their control is accomplished by the transmission of a nerve impulse. A nerve impulse originates in response to a stimulus of an electrical, chemical, thermal, or mechanical nature that has been received by the cell membrane of a neuron. The stimulus elicits a wave of depolarization and repolarization that spreads along the axolemma, away from the site where the stimulus was received, which results in the transmission of the nerve impulse.
Mechanisms of Transmission
The word potential is used in regard to nerve cells as it is in the study of electricity, in which it refers to relative electrical charges between two points in a field or circuit.
For the neuron, this is referred to as a transmembrane potential and the two points are the inside and outside of the confines of the cell membrane. All cells of the body have a transmembrane potential, but the neurons are unique in being able to alter this potential to produce an impulse. The charged transmembrane potential is a local phenomenon close to the cell membrane and does not refer to a charge inside and outside the cell, which is electrically neutral. A measured potential is relatively small, however, and its units are in millivolts rather than volts.Resting Membrane Potential
In a resting neuron, the potential between the two sides of the membrane is called the resting potential. The resting membrane potential results from the unequal distribution of sodium ions (Na+) and potassium ions (K+) on the outside and inside of the neuron. The active transport of Na+ to the outside, coupled with the transport of K+ into the neuron (the Na+-K+ ATPase pump), keeps the concentration of Na+ low on the inside of the membrane. If their rates of transport were equal to each other, electrical neutrality between the inside and outside of the membrane would be maintained. However, the outward active transport of Na+ occurs at a faster rate than the inward active transport of K+, and thus an electronegativity is maintained on the inside of the membrane and an electropositivity on the outside (Figure 4-1q). The membrane is therefore polarized. The resting membrane potential has been measured as about -70 millivolts (mV). It does not exceed -70 mV because, at this level, the electrical gradient is sufficient to cause Na+ diffusion inward to balance the rate of outward active transport.
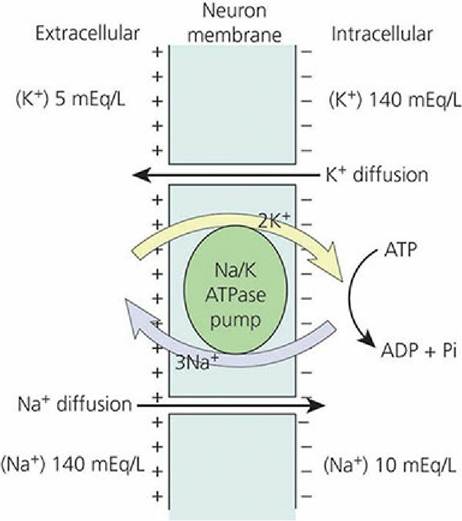
■ FIGURE 4-19 Establishment of a resting membrane potential by active transport of three Na+ outward coupled with the transport of two K+ inward.
The uneven distribution results in the electronegativity within the fiber. The large concentration gradients for sodium and potassium across the resting nerve membrane are caused by the sodium-potassium pump (Na/K ATPase pump). ATP, adenosine triphosphate; ADP, adenosine diphosphate; Pi, inorganic phosphate.Depolarization, Repolarization, and the Nerve Impulse
Chemical or physical stimulation of a neuron increases the permeability of the membrane for Na+ at the point of stimulation and, because there is a high concentration of Na+ on the outside of the membrane in the extracellular fluid, Na+ rushes inward. This reverses the membrane potential at the point of stimulation, so the membrane now becomes positive on the inside and negative on the outside; this is depolarization. The inflow of Na+ soon stops and the permeability of the membrane for K+ increases; the K+ then flows outward because it has a higher concentration inside the neuron than outside. The outflow of K+ reestablishes the resting membrane potential at the point of stimulation; this is repolarization. Measurement of the membrane potential during membrane depolarization and repolarization and its continuous recording on a moving chart is shown in Figure 4-20.
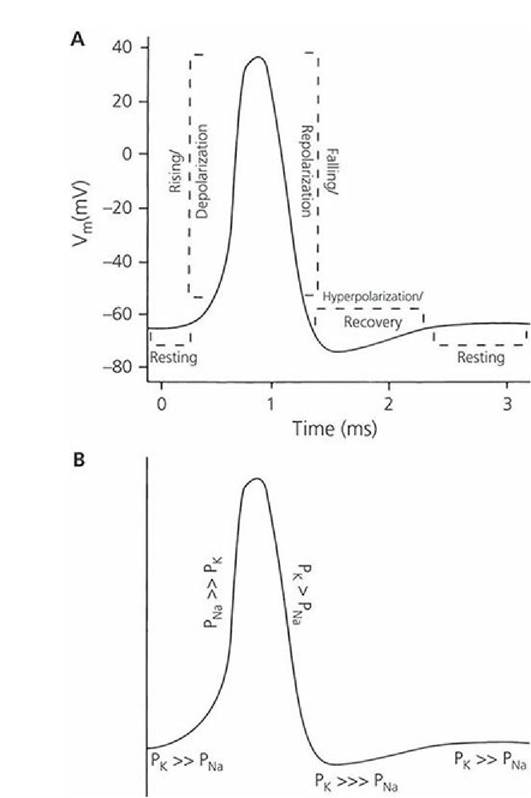
■ FIGURE 4-20 Recording of a transmembrane potential during depolarization and repolarization of a nerve fiber microregion. A. The various phases of the action potential. B. The relative membrane permeability relationships between sodium and potassium ions associated with each of the phases (e.g., PNa >> Pk refers to permeability for Na being greater than permeability for K, where > = greater, >> = much greater, and >>> greatest). Vm, transmembrane voltage; PNa, membrane permeability to sodium; Pk, membrane permeability to potassium.
(From Klein BG. Membrane potentials: the generation and conduction of electrical signals in neurons. In: Reece WO, ed. Dukes’ Physiology of Domestic Animals. 12th edn. Ithaca, NY: Cornell University Press, 2004, 42. Used by permission of the publisher, Cornell University Press.)When a microregion of a nerve fiber is stimulated and subsequently depolarized, a current flow occurs from the point of depolarization to the adjoining microregions. Current flow occurs because a positive charge now exists inside the axolemma at the point of initial depolarization; because of the negative charge inside the axolemma, beyond the point of stimulation, the positive charges (ions) flow toward the negatively charged portion. In addition, the outer aspect of the axolemma (which has become negatively charged at the point of depolarization) attracts positive ions to it from the charged axolemma further ahead. Because of these two events, the interior of the fiber just beyond the depolarized region becomes somewhat more positively charged and the exterior of the fiber just beyond the depolarized region becomes less positively charged. Accordingly, an electrical current flows outward through the axolemma from the interior (that gained positive charges) to the exterior (where positive charges were drawn away). The passage of current out through the axolemma, just beyond the site where depolarization has occurred, causes this region of the membrane to become depolarized in turn (because current flow increases permeability to Na+), just as the axolemma did at the site of the stimulus. The process of depolarization followed by current flow is repeated throughout the length of the nerve fiber and accounts for the nerve impulse (Figure 4-21).
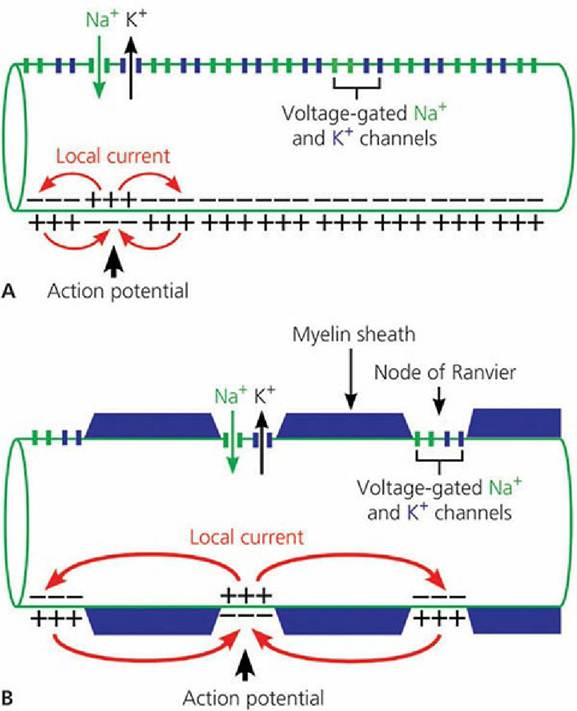
■ FIGURE 4-21 Current paths during the propagation of the action potential in myelinated (A) and nonmyelinated (B) axons. In both axons, the top portion of the membrane illustrates the distribution of the voltage-gated Na+ and K+ channels.
The bottom portion of the axon shows the reversal of membrane polarity triggered by local depolarization. The local currents generated by an action potential flow to adjacent areas of the axonal membrane to depolarize and generate further action potentials. Myelinated axons have Na+ and K+ channels at the node of Ranvier and action potentials jump from one node of Ranvier to the next. This process is referred to as saltatory conduction. (From Uemura EE. Electrochemical basis of neuronal function. In Reece WO, ed. Dukes’ Physiology of Domestic Animals. 13th edn. Wiley-Blackwell, Ames IA, 2015.)Action Potential
Action potentials are changes in the resting membrane potential that are actively propagated along the membrane of the cell. The application of a stimulus to a nerve cell membrane diminishes the resting membrane potential (zero direction). When the membrane potential reaches a critical value (usually 10 to 15 mV less than the resting level of -70 mV), an action potential occurs. The membrane potential at which an action potential is produced is referred to as threshold. Not all stimuli can depolarize the membrane to threshold.
During an action potential, depolarization can change the membrane potential from -70 mV to about +40 mV. During repolarization there is a return to the resting membrane potential of -70 mV. The recording shown in Figure 4-20 represents an action potential. The nerve fiber cannot be stimulated again until repolarization is nearly complete; this is known as the refractory period. When an action potential has been initiated, the nerve fiber is said to fire. If the stimulus is strong enough to initiate an action potential, the entire fiber will fire. This is known as the all-or-none principle for nerve fibers. There is no such thing as a weak impulse. If the stimulus is strong enough to initiate depolarization, the impulse will be conducted with action potentials of normal magnitude. Depolarization and repolarization proceed from one microregion to the adjoining microregion until the entire fiber has been traversed.
Saltatory Conduction
In myelinated fibers the depolarization and repolarization processes are the same, but the action potentials occur from one node of Ranvier to the next instead of over the entire area of the membrane. This process of impulse transmission is referred to as saltatory conduction (saltation refers to an abrupt movement, such as dancing or leaping) (see Figure 4-21). The axolemma is in intimate association with the extracellular fluid at the nodes of Ranvier, and the remainder of the axolemma is relatively insulated from the extracellular fluid..Thus, current flow sufficient to increase membrane permeability leaps from one node of Ranvier to the next, rather than being dissipated at the adjoining microregion. Two functions are served by saltatory conduction. First, impulse transmission is accelerated; second, less membrane is depolarized and repolarized, hence reducing the energy requirement for “recharging” the membrane.
Transmission Velocity
The larger the diameter of the fiber and the thicker the myelin sheath, the faster the transmission of the impulse. The fastest transmission is about 100 m/s and the slowest is about 0.5 m/s. Large myelinated fibers can transmit about 2,500 impulses/s, contrasted with about 250 impulses/s for small unmyelinated fibers.
Final Common Pathway
Somatic lower motor neurons (LMNs) are motor neurons of the spinal cord and brain stem nuclei that innervate skeletal muscle effectors. Upper motor neurons (UMNs) are located in the brain and have fibers that descend to and modify the activity of LMNs. Usually, the branches of many axons (some UMNs), perhaps 2,000 or so, will impinge on the dendritic zone of an LMN, which then, depending on the algebraic sum of the inhibiting and facilitatory input, will either fire or not fire. Thus, the LMN serves as the final common pathway (and the last site for integration) for all output to striated skeletal muscle (Figure 4-22).
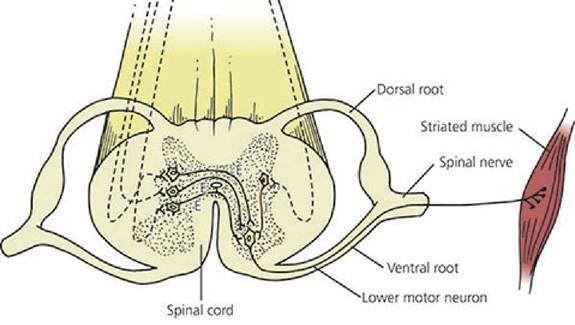
■ FIGURE 4-22 Lower motor neuron going to striated muscle. This represents the final common pathway. To fire, a greater amount of excitatory (E) neurotransmitter must be released than inhibitory (I) neurotransmitter. Dashed lines represent axons of upper motor neurons.
Neuron Placement
Within the central nervous system are several schemes of neuron placement (circuits) that allow for different patterns of activity.
Converging Circuit
A circuit in which several neurons impinge on one neuron is known as a converging circuit (Figure 4-23A). It allows impulses from many different sources to cause some response or provide a sensation.
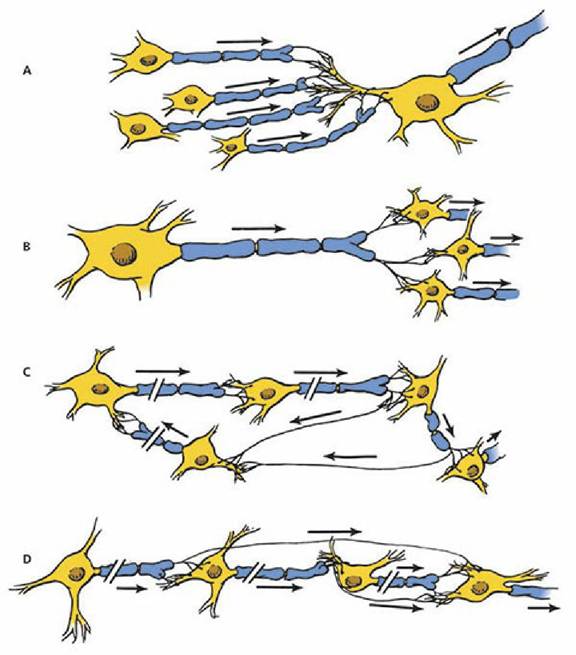
■ FIGURE 4-23 Examples of neuron placement within the central nervous system of mammals. A. Converging circuit. B. Diverging circuit. C. Reverberating circuit. D. Parallel circuit.
Diverging Circuit
A diverging circuit is one in which the axon branches of one neuron impinge on two or more neurons, and each of these in turn impinge on two or more neurons (Figure 4-23B). This type of circuit allows for amplification of impulses and is found in the control of skeletal muscles.
Reverberating Circuit
A reverberating circuit is one in which each neuron in a series sends a branch back to the beginning neuron so that a volley of impulses is received at the final neuron (Figure 4-23C). This type is associated with rhythmic activities, and the volley continues until the synapse fatigues or some other mechanism of unknown type stops the reverberating circuit.
Parallel Circuit
A parallel circuit contains a number,of neurons in a series, with each neuron supplying a branch to the final neuron (Figure 4-23D). Because there is a delay of transmission at the synapse, a volley of stimuli reaches the final neuron. Unlike the reverberating circuit, the impulses then stop. This type provides reinforcement to a single stimulus.
Simple Circuits
Many complex neuron connections are possible, but neuron connections also can be direct and simple. In this regard, the neurons associated with smell (olfactory nerve) and vision (optic nerve) might involve no more than two neurons for their projection to the cerebral cortex. For others, a minimum of three neurons is required to transmit a nerve impulse from the periphery by way of a spinal nerve to the cerebral cortex (Figure 4-24). The three-neuron circuit is the classic circuit for conscious sensations.
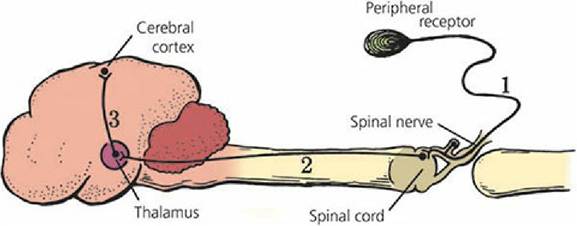
■ FIGURE 4-24 A neuron circuit from periphery to cerebral cortex. A minimum of three neurons is required: (1) afferent neuron in a mixed spinal nerve; (2) neuron ascending in a spinal cord tract to the thalamus; (3) final neuron in the circuit that transmits the impulse to the cerebral cortex.
■