The Skeleton and Joints of the Trunk
The Vertebral Column
The vertebral column (or spine) comprises vertebrae and extends from the skull to the tip of the tail. The vertebrae are firmly but not rigidly joined together.
In addition to contributing to the maintenance of posture through stiffening of the body axis, the spine plays a part in progression and other activities through flexion and extension and sometimes by torsion. The vertebral column encloses and protects the spinal cord and accessory structures within a central canal in particular and shields the structures of the neck, thorax, abdomen, and pelvis in general (see Fig. 2.1).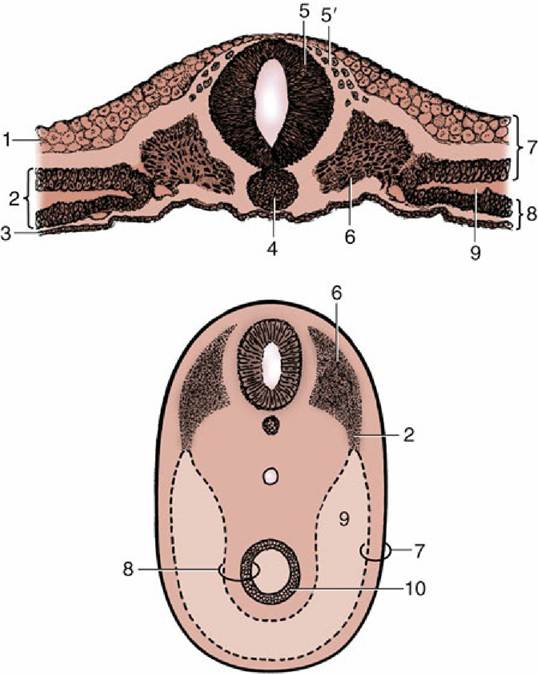
FIG. 2.5 Transections of an early discoidal embryo (top) and of an older ventrally closed one (bottom) to show the splitting of the lateral mesoderm and the development of the celom. 1, Ectoderm; 2, lateral plate of mesoderm; 3, endoderm; 4, notochord; 5, neural tube; 5', neural crest cells; 6, somite; 7, somatopleure;
8, splanchnopleure; 9, celom; 10, primitive gut.
Most vertebrae conform to a common pattern with superimposed features that distinguish the several regions: cervical (C; neck), thoracic (T; back, in the narrow sense), lumbar (L; loins), sacral (S; croup), and caudal (Cd; tail). The numbers of vertebrae that compose these regions vary among species and also, although to a much smaller extent, individually. They can be represented by a formula: that for the dog is C7, T13, L7, S3, Cd20-23.
A typical vertebra consists of a massive body surmounted by an arch that completes the enclosure of a vertebral foramen (Fig. 2.6). The foramina together constitute the vertebral canal. The body, broadly cylindrical, is somewhat flattened on its dorsal surface, which faces into the vertebral canal. It may carry a median crest ventrally.
Its extremities are usually curved: the cranial one is convex, the caudal one concave. The arch consists of two upright pedicles, and from each of these a lamina projects medially to meet its fellow and thus complete the ring about the spinal cord. The bases of the pedicles are notched, and when successive bones articulate, these notches combine to outline intervertebral foramina, openings through which pass both the spinal nerves and the vessels that supply the structures within the vertebral canal. Sometimes an additional lateral vertebral foramen perforates the pedicle next to the intervertebral foramen.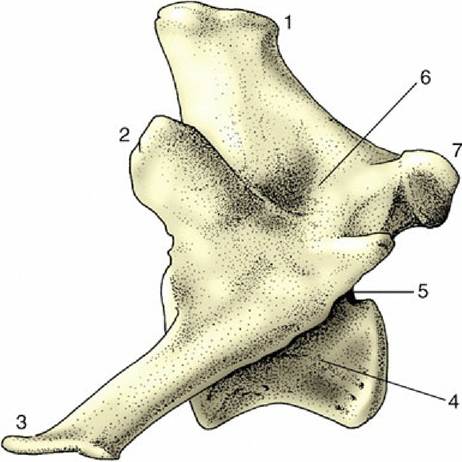
FIG. 2.6 Lumbar vertebra of the dog, left lateral view. 1, Spinous process; 2, cranial articular process; 3, transverse process; 4, body; 5, caudal vertebral notch; 6, arch; 7, caudal articular process.
Each vertebra also carries a number of processes. The generally prominent dorsal or spinous process springs from the union of the laminae, and its length and inclination vary with the region and with the species. Transverse processes arise at the level of the intervertebral foramina and separate the muscles of the trunk into dorsal and ventral divisions. Synovial joints connect restricted parts of the arches. Sometimes the articular facets hardly rise above the level of their surroundings, but elsewhere, and especially in the caudal thoracic and lumbar region, the facets are carried on articular processes that project cranially and caudally from the dorsal portions of the arches (Fig. 2.6/2 and 7).
In domestic as in almost all mammals there are seven cervical vertebrae. The first two, the atlas and the axis, are much modified to allow free movement of the head and require individual description. The remaining five are more typical.
The atlas is the most unusual of all the vertebrae because it appears to possess no body but to consist of two lateral masses joined by dorsal and ventral arches (Fig.
2.7A). This form results from the fusion (in early embryonic life) of a component of the atlantal body with the corresponding part of the following bone, the axis. This addition provides the axis with a cranial projection (dens; Fig. 2.7B/5), which fits into the vertebral foramen of the atlas and serves as a pivot around which the atlas (and the head) may be rotated. A plate of bone, the wing of the atlas (ala atlantis, transverse process), projects laterally from each mass, constituting a landmark that is often visible and always palpable in the living animal. The cranial aspect of the ventral arch and the adjacent areas on the wings carry two deep excavations that receive the occipital condyles of the skull. These facets approach each other ventrally, and in some species they merge. The caudal aspect of the ventral arch is hollowed transversely to articulate with the cranial extremity of the axis. An extension (fovea dentis; Fig. 2.7A/2) of this facet onto the dorsal surface of the ventral arch accommodates the dens. The dorsal arch is perforated by openings that correspond with the transverse and intervertebral foramina of more typical cervical vertebrae; in some species a third (alar) foramen perforates the wing.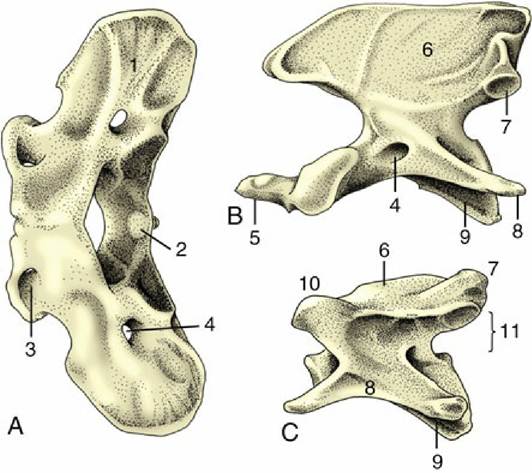
FIG. 2.7 Cervical vertebrae of the dog; cranial is to the left. (A) Atlas, dorsal view. (B) Axis, lateral view.
(C) Fifth vertebra, lateral view. 1, Wing of atlas; 2, fovea dentis; 3, lateral vertebral foramen; 4, transverse foramen; 5, dens; 6, spinous process; 7, caudal articular process; 8, transverse process; 9, body; 10, cranial articular process; 11, position of vertebral foramen.
The axis is the longest vertebra. Its cranial extremity carries the dens, which is rodlike in carnivores and more spoutlike in some other species. The cranial extremity of the body and the ventral surface of the dens together form a single wide articular surface for the atlas.
Dorsally the dens is roughened for the attachment of ligaments that hold it in place. The arch carries a very high (and in the dog, long) spinous process that bears articular facets at its caudal extremity that articulate with corresponding facets on the third cervical vertebra. The transverse processes are large, and each has a transverse foramen in its root to transmit the vertebral artery, vein, and nerve.The remaining cervical vertebrae become progressively shorter as the series is followed toward its junction with the thorax. The extremities of the body are more strongly curved than in other regions and slope obliquely. The ventral surface carries a stout crest. The arch is strong and wide, but the spinous process is poorly developed except on the last (considerable variation, however, exists among species). The large transverse process (Fig. 2.7/8) branches into dorsal and ventral tubercles, the latter commonly developing a caudal platelike extension (Fig. 2.8/5). On the third to sixth bones the process is perforated by a transverse foramen for the passage of the vertebral vessels and nerve. The articular facets are large and flat but do not rise above the surrounding level. The seventh cervical vertebra, transitional to those of the thoracic region, is distinguished by its taller spinous process, its unperforated transverse process, and the presence of facets on the caudal extremity of its body for articulation with the first pair of ribs.
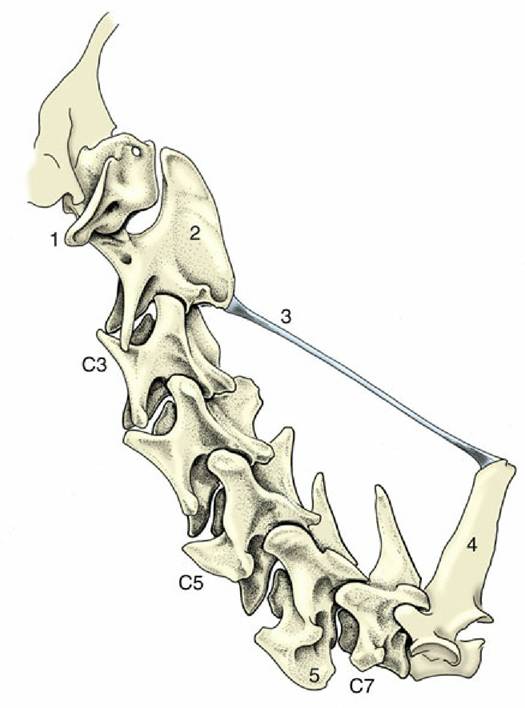
FIG. 2.8 Nuchal ligament of the dog. 1, Wing of atlas; 2, spinous process of axis; 3, nuchal ligament; 4, spinous process of first thoracic vertebra; 5, platelike extension of transverse process.
The thoracic vertebrae (Fig. 2.9) articulate with the ribs and correspond with them in number. Minor variations in number are not uncommon, but the compensation in the lumbar region leaves the thoracolumbar region unaffected.
All thoracic vertebrae have common features, but serial changes also occur that gradually (and on some points abruptly) distinguish the more cranial from the more caudal bones. Common thoracic features are short bodies with flattened extremities; costal facets, on both extremities for the rib heads and on the transverse processes for the rib tubercles; short, stubby transverse processes; closely fitting arches; very prominent spinous processes; and low articular processes.
FIG. 2.9 Thoracic vertebra of the dog, left lateral view. 1, Spinous process; 2, caudal articular process; 3, transverse process with costal fovea; 4, mammillary process; 5, caudal vertebral notch; 6 and 7, costal foveae; 8, body.
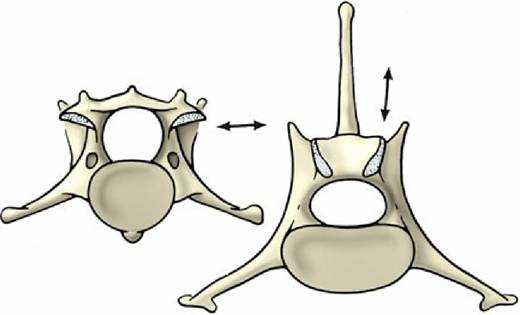
FIG. 2.10 Contrast the orientation (arrows) of the articular surfaces of a cervical vertebra (left) and a lumbar (right) vertebra of the dog, caudal view.
Conspicuous serial features are a rapid increase in the height of the spinous processes, which reaches a maximum a few vertebrae behind the cervicothoracic junction, followed by gradual decline; progressive simplification of the costal facets (those on the transverse processes approach and finally merge with those on the cranial extremity); reduction (and eventual disappearance) of the caudal costal facets; and appearance of an additional (mammillary) process as a projection from the transverse process and its gradual migration to join the cranial articular process. More abrupt changes toward the end of the thoracic series include sudden alteration from a caudodorsal to a craniodorsal orientation of the spinous processes and a change in the character of the articular facets from the cervical to the lumbar pattern (Fig. 2.10). In some species, including the dog, the last members of the thoracic series possess yet other (accessory) processes that spring from the caudal part of the arch to overlap the following bone.
The lumbar vertebrae (Fig. 2.11) differ from the thoracic vertebrae in the greater length and more uniform shape of their bodies. Other regional features are absence of costal facets; a shorter height and generally forward slope of the spinous processes; long, flattened transverse processes that project laterally, sometimes (as in the dog) with a cranioventral inclination; interlocking articular processes; and prominent mammillary, and sometimes also accessory, processes.
Caudal to the loins the vertebral column is continued by the sacrum, a single bone formed by the fusion of several vertebrae. The sacrum forms a firm articulation with the pelvic girdle that transmits the thrust of the hindlimbs to the trunk. Usually only one or two of the constituent vertebrae directly participate in the articulation. The more caudal bones project behind it to furnish the greater part of the roof of the pelvic cavity. In some animals (especially pigs) one or more tail vertebrae may fuse with the sacrum in later life. In the dog the three sacral vertebrae form a short quadrilateral block (Fig. 2.12).
The sacrum commonly narrows from its cranial to its caudal extremity and is curved along its length to present a smooth, slightly concave face to the pelvic cavity. In most species the dorsal surface is marked by the appropriate number of spinous processes, although they may be much reduced or even absent (e.g., pig). They may preserve their independence (e.g., dog or horse) or fuse to form a continuous crest (e.g., ruminants). Lateral to this crest, a lower irregular crest usually marks the site of the redundant articular processes. The margin of the bone is formed by the fused transverse processes and carries toward its cranial extremity the articular surface for the ilium; this is often ear-shaped, hence the name auricular surface (Fig. 2.12/2).
The degree of fusion, least in the pig, of the sacral vertebrae varies among species. Even when fusion is total, the composition of the sacrum is betrayed by the number of foramina that mark both surfaces; the dorsal and the ventral branches of the sacral nerves issue separately through them. The junction of the ventral surface with the cranial extremity forms a lip known as the promontory (Fig. 2.12/1); though often inconspicuous, it is a reference point in obstetrics.
The number of caudal vertebrae varies greatly, even within a single species. These vertebrae show a progressive simplification in form, and although the first few resemble miniature lumbar vertebrae, the middle and later members of the series are reduced to simple rods. In addition to the usual features, the more cranial vertebrae of some species provide protection to the main artery of the tail in the form of ventral (hemal) arches, separate small chevron (V-shaped) bones connected to the undersurfaces of the bodies, or paired ventral (hemal) processes (Fig. 2.12E).
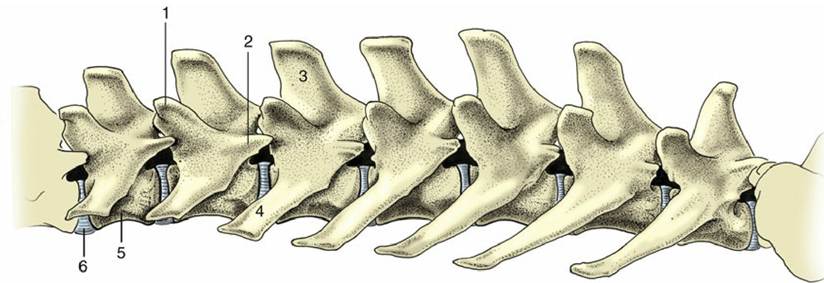
FIG. 2.11 Lumbar vertebrae of the dog, left lateral view. 1, Mammillary process; 2, accessory process; 3, spinous process; 4, transverse process; 5, body; 6, intervertebral disk.

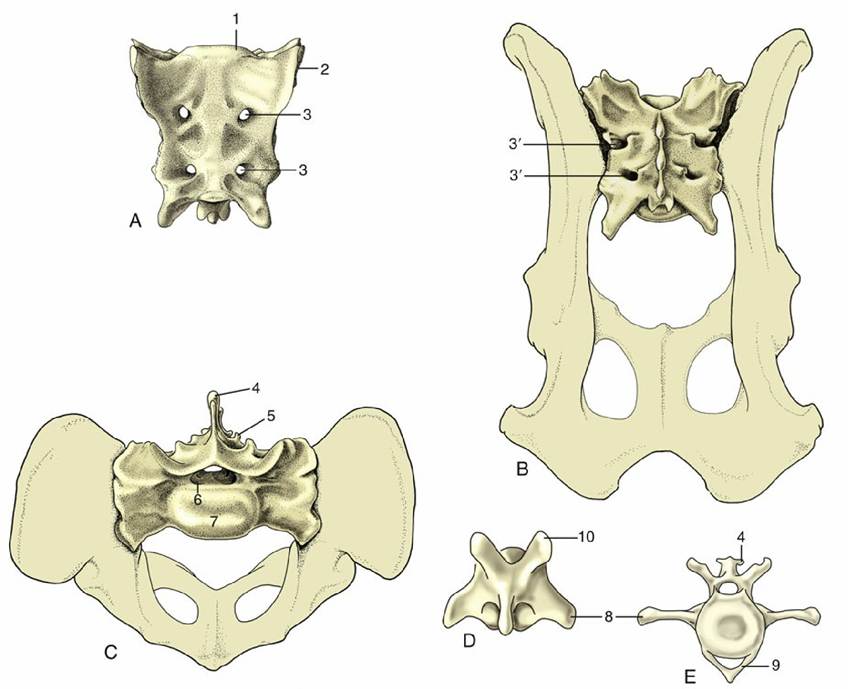
FIG. 2.12 Canine sacrum and caudal vertebrae. (A) Sacrum, ventral view. (B) Sacrum, dorsal view. (C) Sacrum, cranial view. (D) Caudal vertebra, dorsal view. (E) Caudal vertebra, cranial view. 1, Promontory; 2, auricular articular surface; 3, ventral (3' dorsal) sacral foramina for ventral (3' dorsal) branches of sacral nerves; 4, spinous process; 5, rudimentary articular process; 6, vertebral canal; 7, body; 8, transverse process; 9, hemal arch, also called chevron; 10, cranial articular process.
The contours of the vertebral column vary with the posture, the species, and the breed. In general, the vertebrae from the caudal thoracic region to the tail head follow a more or less horizontal line. The more cranial thoracic vertebrae slope downward to reach the lowest point at the entrance to the chest, where an abrupt change in direction puts the spine on a course that ascends toward the head. The ventral inclination of the cranial thoracic vertebrae is masked in the live animal by the height of the spinous processes; indeed, in some species, the horse most notably, the spines are so long that the contour of this part of the back is raised to constitute the withers. Except toward the poll, the cervical vertebrae run at some distance from the dorsal skin. This feature is not apparent in the live subject, and in larger animals it may not be easy to determine, even on palpation. The greater part of the tail hangs down in large animals, but its posture is more variable in dogs and cats, being an expression of emotion in both species and influenced by breed in the former.
The Joints of the Vertebral Column
The vertebrae form two sets of joints: one cartilaginous, involving the direct connection of the vertebral bodies, the other synovial, existing between facets carried on the vertebral arches. In addition, certain long ligaments extend over many vertebrae. This pattern is modified in two regions; cranially, allowance is made for the free movement of the head, and in the pelvic region, sacral fusion occurs.
The two joints of the atlas are described first. The atlanto-occipital joint (Fig. 2.13) is formed between the condyles of the skull and the corresponding concavities of the atlas. Although the separate right and left articular surfaces converge ventrally, they do not always merge, although generally there is a single synovial cavity. The synovial membrane attaches around the occipital and atlantal facets. It is strengthened externally by dorsal and ventral atlantooccipital membranes, which pass from the arches of the atlas to corresponding parts of the margin of the foramen magnum (see Fig. 2.32/12), and by lesser lateral ligaments, which pass between the atlas and adjacent regions of the skull. Despite its odd character, the joint functions as a ginglymus: movement is virtually restricted to flexion and extension in the sagittal plane (the nodding movement in humans).
The atlantoaxial joint is even more peculiar. The extensive articular surfaces of the ventral arch of the atlas and of the body and dens of the axis face into a single synovial cavity. The surfaces are so formed that only limited areas are in contact in any position of the head. This limitation of contact, together with the roomy capsule, allows some versatility of movement, although mainly confined to rotation about a longitudinal axis (the head-shaking movement in humans). The dorsal atlantoaxial ligament that joins adjacent parts of these vertebrae imposes little restraint.
The Axis and Death: The dens of the axis is strapped by one or more ligaments to the adjacent part of the upper surface of the ventral atlantal arch and sometimes also to the occipital bone (as in the dog). In judicial hanging, the rupture of these ligaments — or fracture of the dens itself— allows the axis to strike against the cord and cause death.
A single description serves for the articulations of most other vertebrae. The intervertebral articulations combine symphyses between the bodies and synovial joints between the articular processes. The thick flexible pads, the intervertebral disks, connect the bodies of adjacent vertebrae and contribute to about 10%, 16%, and 25% of the vertebral length in ungulates, dogs, and humans, respectively. The disks are among the organs that most consistently show degenerative changes with advancing age and are a common source of back troubles long recognized in ourselves and in dogs, which are now also diagnosed in other domestic and even wild animals. But the nature of the medical conditions related to disks is not the same in humans as in quadrupeds.
Each disk consists of two parts, a nucleus pulposus and an anulus fibrosus (Fig. 2.14). The nucleus occupies a slightly eccentric position. In the young animal, it consists of an unusual semifluid tissue derived from the embryonic notochord and retains some resemblance to it in structure. It is contained under pressure and escapes if afforded opportunity. The anulus fibrosus consists of encircling bundles of fibrous tissue that pass obliquely from one vertebra to the other, in most species merging with cartilage plates that cap the bones. The orientation of the fibers changes between about 20 successive lamellae. The distinction between anulus and nucleus is not always very clear, particularly in the larger species. Retention of the nucleus within the fibrous ring absorbs shock and spreads the compressive forces to which the column is subjected over a wider part of the vertebrae.
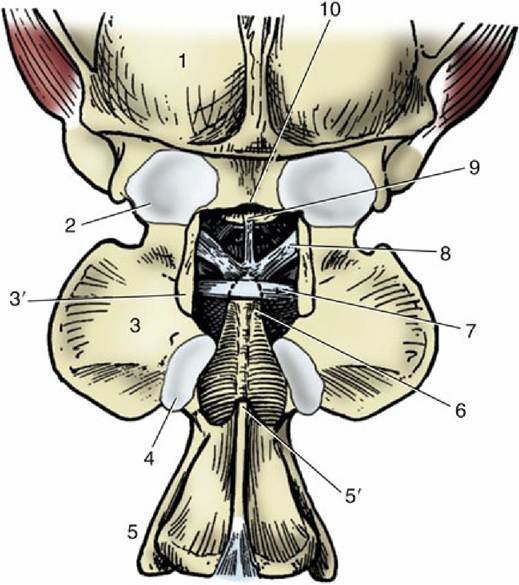
FIG. 2.13 Canine atlanto-occipital joint, dorsal view; the dorsal arch of the atlas has been removed. 1, Skull; 2, atlanto-occipital joint capsule; 3, wing of atlas; 3', dorsal arch of atlas, resected; 4, atlantoaxial joint capsule; 5, axis; 5', spine of axis, its overhanging cranial portion having been removed; 6, dens; 7, transverse ligament of atlas; 8, alar ligaments; 9, apical ligament of dens; 10, dorsal margin of foramen magnum.
The Intervertebral Disk Pathology: Both nucleus and anulus start to change in a subtle manner rather early in life. For example, fragmentation of the ring leads to escape of the nucleus into the vertebral canal and subsequent pressure on the spinal cord. Age-related calcification of the nucleus reduces the normal resilience and flexibility of the spine. Degenerative changes may affect any disk, but the disks at the most mobile regions, such as the neck and, in large animals, the lumbosacral junction, are especially susceptible. Most thoracic disks are crossed dorsally by the intercapital ligaments that unite the heads of the right and left ribs (p. 39), and these ligaments are alleged to mitigate the effects of disk rupture at these levels.
The synovial joints are interposed between the vertebral arches, and the joint mobility varies with the spaciousness of the joint, the region, and to some extent the species. In the cervical and cranial thoracic regions the joint surfaces are arranged tangential to the circumference of a circle centered in the vertebral body (see Fig. 2.10), which allows rotation in addition to flexion and extension. The radial alignment of the articulating surfaces in the caudal thoracic and lumbar regions restricts the movement mostly to the median plane. Movement is most free in the neck, where the articular surfaces are largest and the capsules loosest. The elastic interarcuate ligaments fill the dorsal spaces between the arches of successive vertebrae, and their extent is inversely related to the width of the arches. In certain regions, interspinous and intertransverse ligaments also exist, but these are of less importance.
Three long ligaments extend along substantial portions of the column. A dorsal longitudinal ligament (Fig. 2.15/7) runs along the floor of the vertebral canal from the axis to the sacrum, narrowing over the middle of each vertebral body and widening where it crosses each intervertebral disk. A ventral longitudinal ligament follows the ventral aspect of the vertebrae from the midthoracic region to the sacrum; more cranially, its role is filled by the longus colli muscles. It also widens over and fuses with the intervertebral disks.
A third (supraspinous) common ligament runs over (or to each side of) the summits of the spinous processes of the thoracic and lumbar vertebrae. It merges with the tendons of the epaxial muscles so completely that some writers dispute its independent existence. Except in the pig and cat, a cranial continuation of this ligament leaves the highest spines of the withers and runs by the shortest route to attach to the nuchal surface of the skull or, as in the dog, the spinous process of the axis (see Fig. 2.8). This nuchal ligament runs close to the upper contour of the neck, and for most of its length, it is well separated from the more ventral course followed by the cervical vertebrae. Unlike the other long ligaments, this ligament is elastic and supports the burden of the head when it is held high without interfering with the animal's ability to lower the head to feed or drink from the ground. There is an obvious correlation between the strength of this ligament and the weight of the head and the length of the lever arm of the neck; the nuchal ligament is therefore much more powerfully developed, and more complicated, in the larger species (see Fig. 19.3).
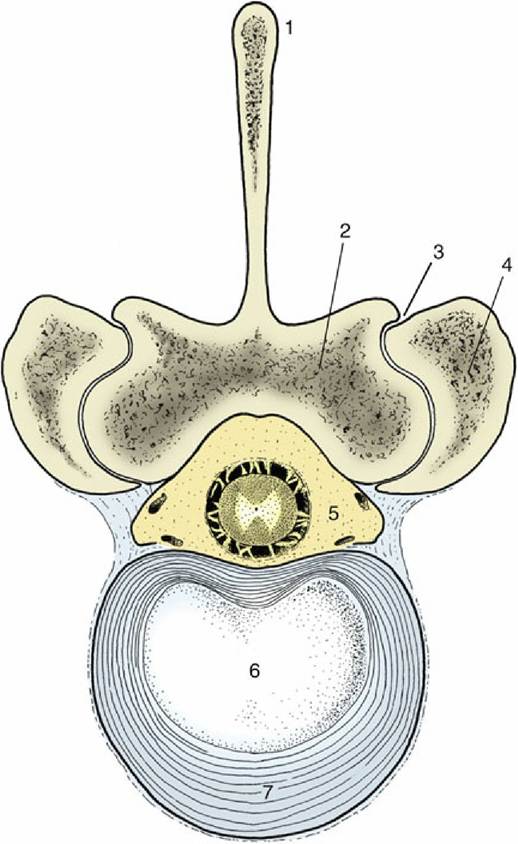
FIG. 2.14 Bovine lumbar intervertebral disk. 1, Spinous process; 2, lamina; 3, synovial intervertebral joint; 4, articular process of adjacent vertebra; 5, vertebral canal with contents (spinal cord and meninges surrounded by epidural fat); 6, nucleus pulposus; 7, anulus fibrosus.
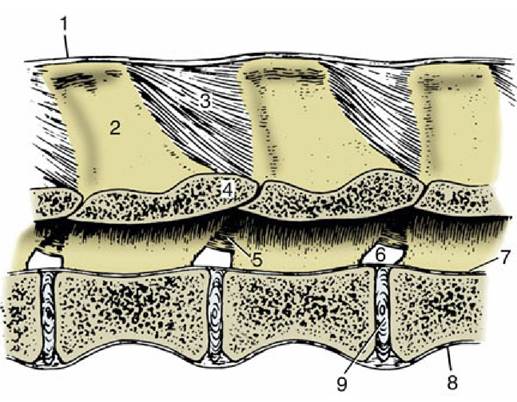
FIG. 2.15 Ligaments of the vertebral column. Paramedian section of lumbar vertebrae of a dog, viewed from the left. 1, Supraspinous ligament; 2, spinous process; 3, interspinous ligament; 4, arch of vertebra; 5, interarcuate ligament; 6, intervertebral foramen; 7, dorsal longitudinal ligament; 8, ventral longitudinal ligament; 9, intervertebral disk.
The Ribs and Sternum
The thoracic skeleton is completed by the ribs and sternum. The ribs (costae) are arranged in pairs and generally articulate with two successive vertebrae: the caudal vertebra is the one with the same numerical designation as the rib. Each rib consists of a bony dorsal part, the rib proper, and a cartilaginous ventral part, the costal cartilage (Fig. 2.16A). The two parts meet at a costochondral junction. The dorsal part of the rib articulates with the vertebral column, whereas the cartilage articulates with the sternum either directly, as do the first eight or so sternal or "true" ribs, or indirectly through connection of the cartilage with that in front, as do the asternal or "false" ribs. In this way, the cartilages of the asternal ribs combine to form the costal arch (Fig. 2.17A/6), the cranial boundary of the flank. The cartilage of the last rib may fail to make contact with its neighbor, and this rib is then said to be "floating."
The dorsal extremity of the rib terminates in a rounded head that carries two facets, one for articulation with the body of each of the two vertebrae with which it is connected. These facets are separated by a rougher area (crest) that makes contact with the intervertebral disk and on most ribs also gives origin to the intercapital ligament. The head is joined to the body of the rib by a short constricted neck whose lower part carries a lateral tubercle. The tubercle bears a third articular facet, which meets that on the transverse process of the more caudal of the associated vertebrae (Fig. 2.16B).
The body of the rib begins beyond the tubercle. It is long, curved in its length, and usually laterally flattened, particularly in the larger species and toward the lower end. It is most strongly bent at a region known as the angle (Fig. 2.16/4), where the lateral surface is roughened for the attachment of the iliocostalis. The cranial and caudal margins of the body are often sharply defined and give attachment to the intercostal muscles that fill the space between successive ribs. The caudal margin may also be grooved to give protection to the neurovascular bundle of the intercostal space.
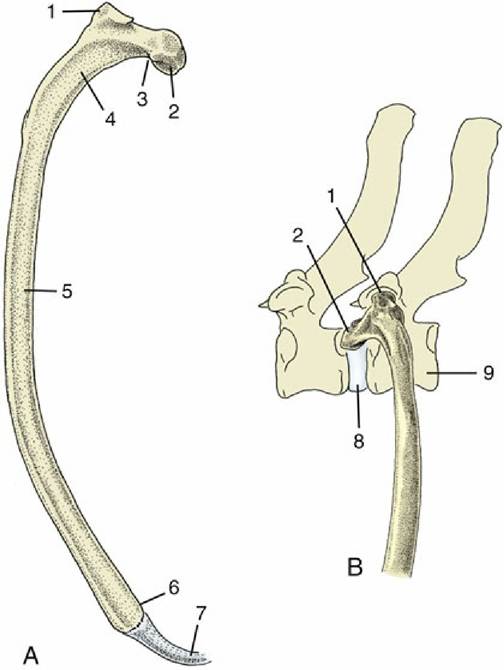
FIG. 2.16 (A) Left rib of a dog, caudal view. (B) Left rib of a dog articulating with two vertebrae, lateral view. 1, Tubercle; 2, head; 3, neck; 4, angle; 5, body; 6, costochondral junction; 7, costal cartilage; 8, intervertebral disk; 9, vertebra of same number as rib.
The costal cartilage is flexible in the young animal, especially if it is long and thin, as in the dog. It becomes more rigid as calcification develops and increases with age. The cartilage either meets the bony rib at an angle (knee, genu) or is itself flexed cranioventrally some way beyond the costochondral junction.
Serial changes are obvious. The first rib is always relatively strong, short, and straight. Its cartilage is also stumpy and articulates with the sternum at a tight joint that fixes the rib to create a firm base toward which the other ribs may be drawn on inspiration. The succeeding ribs increase in length, in curvature, and in caudoventral inclination, most markedly over the caudal part of the thoracic wall, although the very last two or three may be somewhat shorter. The three articular facets of the upper end approach and eventually merge on the ribs toward the end of the series. The cartilages of the sternal ribs are short and about as thick as the bony ribs; those of the asternal ribs are mostly slender and taper toward their ventral extremities.
The sternum is composed of three parts. The most cranial part, known as the manubrium (Fig. 2.17/1), generally projects in front of the first ribs and may be palpated at the root of the neck. It is rodlike in the dog and cat but is laterally compressed in the larger animals. The body of the bone is composed of several segments (sternebrae), in youth joined by cartilage that later ossifies. The sternum is cylindrical in the dog, is wide and flat in ruminants, and carries a ventral keel in the horse (Fig. 2.17B). Its dorsolateral margin bears a series of depressions that articulate with costal cartilages. The more cranial of these depressions alternate with the sternebrae, and each receives a single cartilage; the more caudal depressions are crowded more closely together and may receive more than one cartilage. The caudal part of the sternum consists of flat (xiphoid) cartilage (Fig. 2.17/5) that projects between the lower parts of the costal arches. It supports the most cranial part of the abdominal floor and gives attachment to the linea alba.
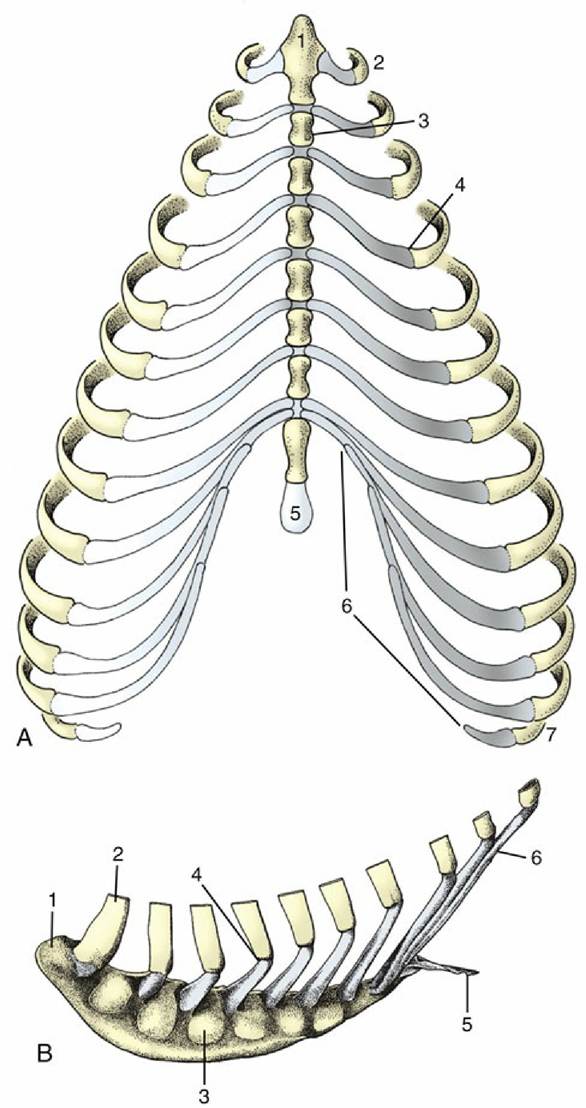
FIG. 2.17 (A) Canine and (B) equine sternum and costal cartilages, ventral and left lateral views. 1, Manubrium; 2, first rib; 3, sternebra; 4, costochondral junction; 5, xiphoid cartilage; 6, costal arch; 7, floating rib.
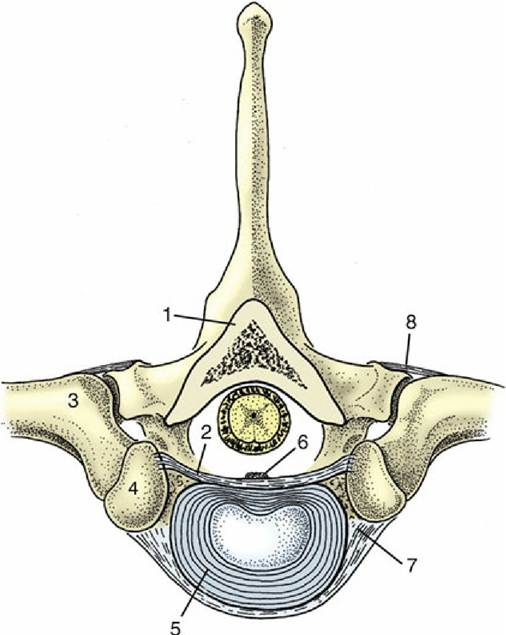
FIG. 2.18 Costovertebral articulations; transverse section of the vertebral column of the dog (about T8). 1, Lamina of vertebra; 2, intercapital ligament; 3, tubercle of rib; 4, head of rib; 5, intervertebral disk; 6, dorsal longitudinal ligament; 7, costovertebral joint; 8, costotransverse joint covered by costotransverse ligament.
The Joints of the Thoracic Wall
Most ribs make two separate articulations with the vertebral column. The head participates in a ball-and-socket costovertebral joint of unusually restricted mobility. The joint cavity is divided into two compartments by the intercapital ligament (Fig. 2.18/2), which arises from the interarticular crest. This ligament passes through the intervertebral foramen, crosses the floor of the vertebral canal, and ends by inserting on the corresponding region of the rib of the other side. In its passage, it detaches slips that anchor to the intervertebral disk and the adjacent parts of the vertebrae. It passes below the dorsal longitudinal ligament (Fig. 2.18/6) and offers some protection against nuclear material from protrusion of a ruptured disk into the vertebral canal. An intercapital ligament is not found at the first costovertebral joint or at the last few. Additional short and tight ligaments support the joint dorsally and ventrally.
The costotransverse joint in which the tubercle participates is of the sliding variety. It is supported by a ligament that passes between the neck of the rib and the transverse process of the vertebra (Fig. 2.18/8).
The costosternal joints are synovial joints of the pivot variety. The interchondral joints of the asternal ribs are syndesmoses of a rather elastic nature. The intersternal joints are mostly impermanent synchondroses, although in some species the manubrium articulates with the body at a synovial joint.
The movements possible at these joints are discussed with the actions of the muscles of the thoracic wall.
The Pelvic Girdle
Although the pelvic girdle is formally a part of the hindlimb skeleton, discussing it here seems more sensible because it is fully integrated into the construction of the trunk. The girdle consists of symmetrical halves, the hip bones (ossa coxarum), which meet at the pelvic symphysis ventrally and form firm, though not rigid, articulations with the sacrum dorsally. When augmented by the sacrum and first few tail vertebrae, the girdle forms a ring known as the bony pelvis around the pelvic cavity. The form of the bony pelvis reflects a compromise between physiologic processes, of which those related to giving birth are most important, and the requirements of locomotion and posture.
Each hip bone is composed of three bones that develop from separate ossifications within a single cartilage plate. The strips of cartilage that demarcate the boundaries between the bones to allow for growth disappear once growth is complete. It is therefore artificial to describe the three components — ilium, pubis, and ischium—as separate units. The ilium (Fig. 2.19/1) is the craniodorsal part that extends obliquely forward from the hip joint to articulate with the sacrum. The pubis (Fig. 2.19/6) extends medially from the joint to form the cranial part of the pelvic floor. The ischium (Fig. 2.19/8) is more caudal and forms the larger part of the floor, although it also sends a branch to the joint. Both pubis and ischium participate in the symphysial joint in domestic species, although only the pubis does so in the human pelvis.
The ilium consists of a cranial expansion or wing and a caudal shaft or body. The wing is oblong with a more or less sagittal orientation in the dog and cat but is triangular and almost vertical in the horse and ruminants (see Fig. 2.19). Its margin forms saliences, generally thickened, at certain points. Dorsally (dorsomedially in the larger species), it forms a sacral tuber that is reduced to two low (cranial and caudal dorsal iliac) spines in the dog and cat (Fig. 2.19/3) but is prominent in the large animals, in which it is close to the spinous processes of the vertebrae (Fig. 2.19/3'). Ventrally (ventrolaterally in the larger species), the ilium forms a coxal tuber (Fig. 2.19/2' and 2) which is also reduced to low (cranial and caudal ventral iliac) spines in the carnivores but is prominent in large species, forming the point of the hip at the dorsocaudal corner of the flank (Fig. 2.20B/8). Including these projections, the margin of the wing is known as the iliac crest; thickened and convex in carnivores, it is thin and concave in large animals. Some of these features form important landmarks in the living animal.
The lateral (dorsolateral) surface is excavated and largely given over to the origin of the gluteus medius, whose attachment may raise one or more quite prominent ridges. The medial (ventromedial) surface faces toward the body cavity. The ventral part gives origin to the iliacus, whereas more dorsally it bears the roughened auricular articular surface for the sacrum (see Fig. 2.19B/15). The dorsal border of the wing at its junction with the shaft forms the greater sciatic notch (incisura; see Fig. 2.19/4), over which the sciatic nerve runs in passage to the hindlimb.
The shaft of the ilium is robust and columnar. Its caudal extremity contributes to the acetabulum, the deep cavity that receives the head of the femur. Its ventral border is marked by the low arcuate line that serves as part of the arbitrary boundary ("terminal line") between the abdominal and pelvic cavities. Except in the dog, the line carries the psoas tubercle midway along its length, where the psoas minor attaches.
The pubis (Fig. 2.19/6), essentially L-shaped, consists of cranial (acetabular) and caudal (symphysial) branches. The lateral end of the cranial branch contributes to the acetabulum and is known as the body. Its cranial edge, known as the pecten of the pubis, bears the iliopubic eminence and gives attachment to the abdominal muscles. Between them, the two branches account for about half the circumference of the obturator foramen (Fig. 2.19/7), the large opening in the pelvic floor through which the obturator nerve emerges. The foramen is closed by muscle and membrane in the fresh state.
The ischium (Fig. 2.19/8) is a horizontal plate extended cranially by symphysial and acetabular branches, one to each side of the obturator foramen. The extremity of the acetabular branch that contributes to the articular cup is known as the body. The body and the cranial part of this branch are surmounted by a crest, the ischial spine (Fig. 2.19/5), which also extends onto the caudal part of the ilium. Marked by the origin of the gluteus profundus, ischial spine is relatively low in the dog and particularly high in ruminants. The caudolateral corner of the plate forms the ischial tuber (Fig. 2.19/9); the border between this tuber and the spine is indented by the lesser sciatic notch (Fig. 2.19/10). The ischial tuber is a horizontal thickening in the dog and a conspicuously triangular swelling in cattle. In most species it is subcutaneous, and it may be a visible landmark. The remaining part of the caudal border forms with its fellow the ischial arch, a notch that is broad and, except in the horse, shallow.
The acetabulum is a deep articular cup to which all three bones contribute; an additional small acetabular bone may be found in young animals. The acetabulum is contained by a prominent rim that is interrupted by a notch caudoventrally. It carries a lunate articular surface internally, but the depth of the cup is nonarticular and rough.
The form of pelvic girdle differs among species. The larger and heavier species have a nearly vertical ilium that brings the sacroiliac joint, and therefore the weight of the trunk, more nearly above the hip joint (see Fig. 2.20B). In smaller species, the highly oblique ilium (see Fig. 2.1) displaces the pelvic floor caudally relative to the vertebral column and increases the effectiveness of the abdominal muscles that flex the column in bounding gaits. Caudal displacement of the ischial tuber also increases the leverage that may be exerted by the hamstring muscles that arise here.
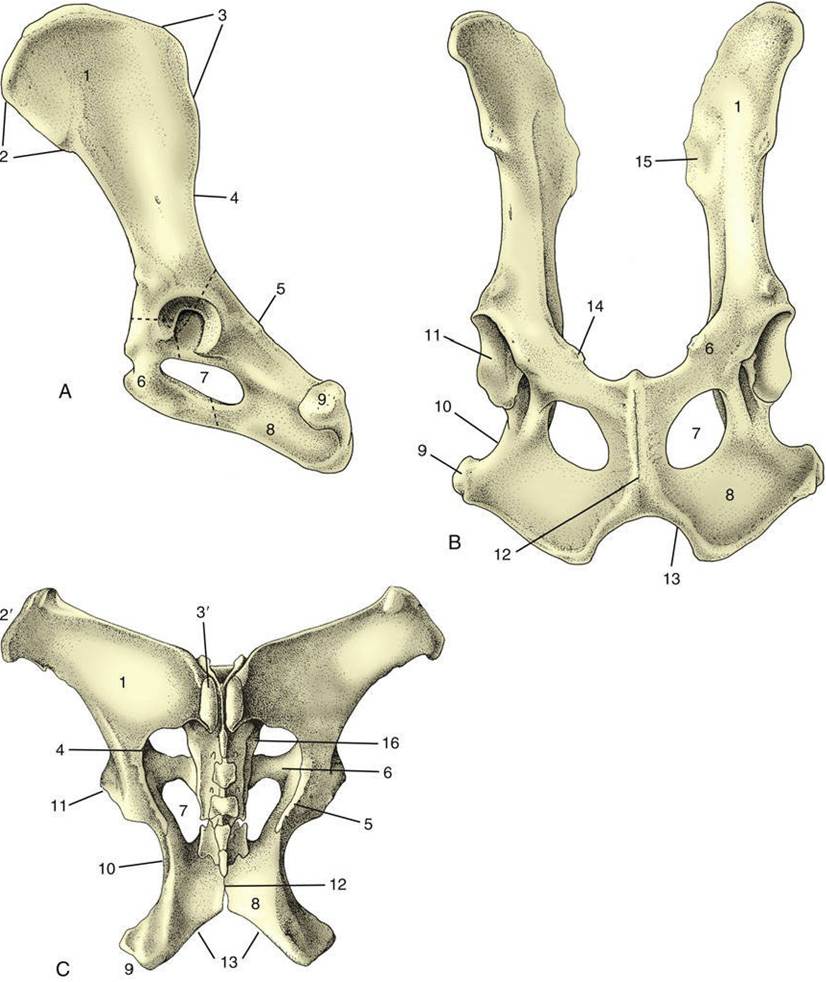
FIG. 2.19 Canine hip bones in left lateral (A) and ventral (B) views. (C) Dorsal view of equine pelvis. The broken lines give the approximate extents of ilium, pubis, and ischium. 1, Wing of ilium; 2, ventral iliac spines; 2', coxal tuber; 3, dorsal iliac spines; 3', sacral tuber; 4, greater sciatic notch; 5, ischial spine; 6, pubis; 7, obturator foramen; 8, ischium; 9, ischial tuber; 10, lesser sciatic notch; 11, acetabulum; 12, pelvic
symphysis; 13, ischial arch; 14, iliopubic eminence; 15, auricular articular surface; 16, sacrum.
The dimensions of the girdle are most important in species that carry a single large offspring. They are of little significance in polytocous species (those that normally carry a litter), in which the full-term fetuses are relatively small. These aspects of pelvic conformation are discussed in later chapters.
The Joints and Ligaments of the Pelvic Girdle
The pelvic symphysis is a secondary cartilaginous joint that ossifies with advancing age. The process of ossification is irregular because it commences at different ages and advances at different rates, even in a single species. It usually starts earlier and is more advanced at any stage in the pubic than in the ischial part. It is sometimes asserted that in certain domestic species changes can be detected in the tissues of the symphysis (and sacroiliac joint) in advance of parturition. These changes, if present, are minor in comparison with those that occur in guinea pigs and many other small animals at this time; in the latter animals, complete dissolution of the symphysis, which allows the two halves of the girdle to move apart to enlarge the birth passage, may occur.
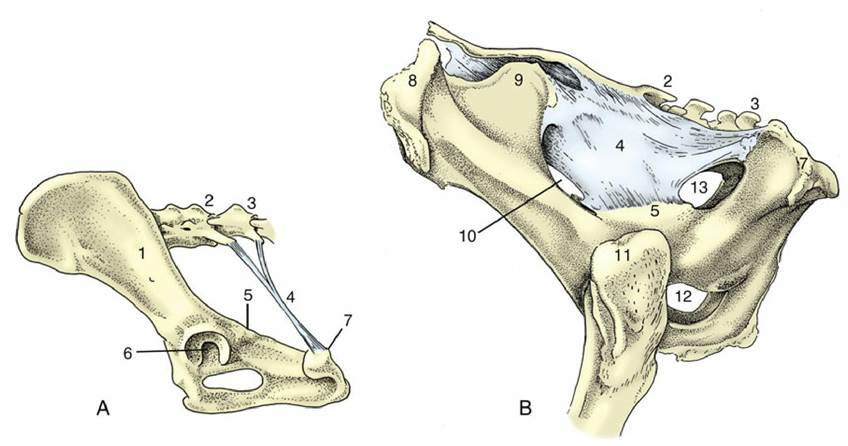
FIG. 2.20 Canine sacrotuberous ligament (A) and bovine sacrosciatic ligament (B), left lateral views. 1, Ilium; 2, sacrum; 3, caudal vertebra(e); 4, sacrotuberous ligament in A, sacrosciatic ligament in B; 5, ischial spine; 6, acetabulum; 7, ischial tuber; 8, coxal tuber; 9, sacral tuber; 10, greater sciatic foramen;
11, greater trochanter; 12, obturator foramen; 13, lesser sciatic foramen.
The sacroiliac joints are curious in combining a synovial joint with an adjacent region of extensive fibrous union. The arrangement appears designed to combine firmness of attachment with some shock-absorbent capacity, because these joints are required to transmit the weight of the trunk to the hindlimbs during standing and the thrust of the limbs to the trunk during progression. The sacrum is wedged between the two halves of the pelvic girdle. Each sacral wing carries an articular surface that is broadly flat (but irregular in detail) to match the corresponding iliac surface. The joint capsule is tight and is surrounded and supported by short fascicles of connective tissue that join adjacent parts of the two bones. It is a matter of preference whether certain longer sacroiliac ligaments, at a greater distance from the synovial articulation, are to be regarded as components of that joint or as independent structures. They may include long and short dorsal ligaments passing between the wing of the ilium and the spinous processes and other features of the sacrum. A ventral ligament offers more immediate support to the joint.
The sacrotuberous ligament (Fig. 2.20/4) is of considerably greater interest. In the dog, it is a stout rounded cord extending between the caudolateral angle of the sacrum and the lateral part of the ischial tuber; no such ligament is present in the cat. In ungulates, it is better named the sacrosciatic ligament because it is expanded to a broad sheet that largely fills the space between the lateral border of the sacrum and the dorsal border of the ilium and ischium, which leaves open two foramina adjacent to the greater and lesser sciatic notches. The caudal edge is palpable in dogs and cattle (see p. 481 and p. 686).