The stomach is the dilated part of the digestive tract where digestion is initiated.
It is succeeded by the intestine, which consists of a proximal small intestine (the principal organ of digestion and absorption in most species) and a distal large intestine (generally much shorter and especially concerned with the dehydration of the food residue).
There is considerable diversity, much of it due to dietary habits, among mammals in the form and function of stomach and intestines, which are collectively known as the gastrointestinal tract (Figs. 3.35 and 3.36). The concentrated diet of carnivores is most easily digested, and these animals have a small and simple stomach (Fig. 3.35A) and a relatively short and uncomplicated intestine. The fodder of herbivores has a lower nutritive value and must be consumed in large amounts. Moreover, celluloses and other complex carbohydrates, major parts of the diet, need to be broken down by symbiotic microorganisms for their enzymatic breakdown and utilization. This relatively slow process requires the provision of a large fermentation chamber to create an environment favorable to the multiplication and activity of the microorganisms. Such a chamber is supplied by a greatly enlarged and subdivided stomach (e.g., ruminants) or by a voluminous and complicated large intestine (e.g., horses). Species-specific details are found later in the book, and the following description is largely confined to the dog and cat.
The stomach (ventriculus)* receives food from the esophagus and retains it for a time before discharging it into the duodenum, the first part of the small intestine. The stomach of the dog has a relatively modest capacity, ranging from 0.5 to 6.0 L according to breed, and conforms to a pattern that is common to most carnivores and indeed to many other mammals, including humans. It consists of two distinct parts that converge and join at a ventral angle (Fig. 3.37). The larger part, into which the esophagus opens at the cardia, lies mainly to the left of the median plane, well forward under cover of the ribs and in direct contact with the liver and the diaphragm.
The cardia is relatively distensible and rapidly expands to accommodate a meal. The second part is narrower, has thicker walls, and is more constant in appearance because it is less affected by the presence of a meal. It passes to the right to continue into the duodenum at the pylorus (Fig. 3.35B). The cranial (parietal) aspects of both parts are mainly in contact with the liver, whereas the more numerous relations of the caudal (visceral) surface include the intestinal mass, left kidney, pancreas, and greater omentum. The left part of the margin is applied to the hilar region of the spleen.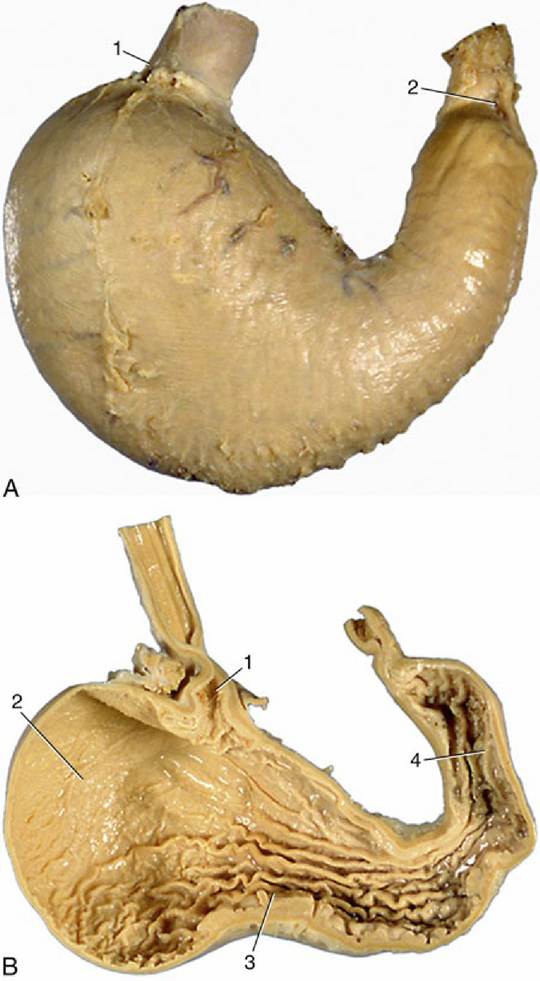
FIG. 3.35 (A) Visceral surface of stomach (dog). 1, Cardia; 2, pylorus. (B) Interior of stomach (dog). 1, Cardiac opening; 2, fundus; 3, body; 4, pyloric antrum.
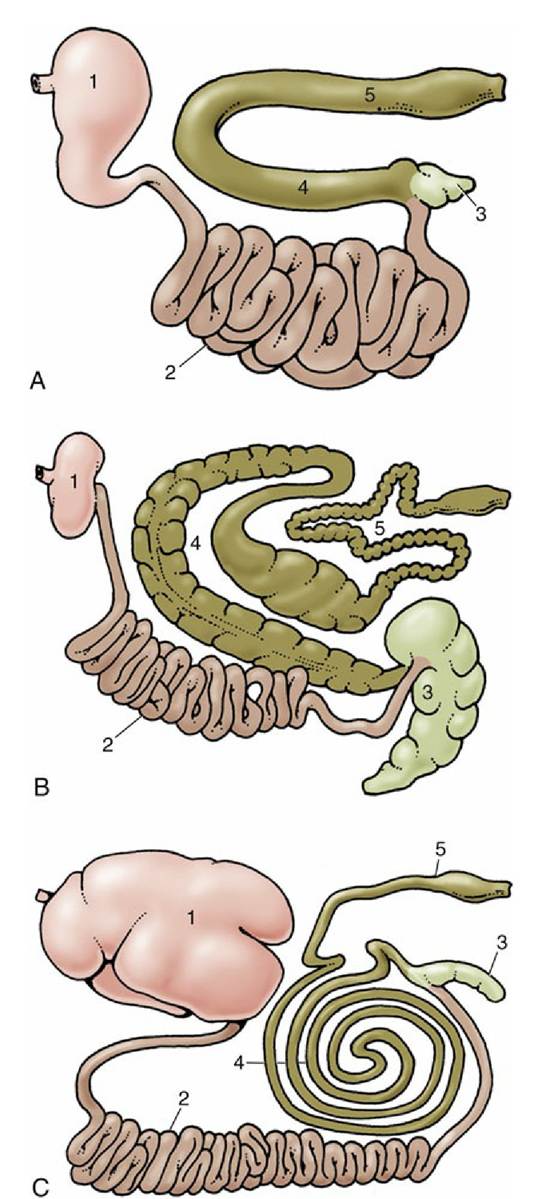
FIG. 3.36 Gastrointestinal tracts of (A) the dog, (B) the horse, and (C) cattle laid out in one plane. 1, Stomach; 2, small intestine; 3, cecum; 4, ascending colon; 5, descending colon.
The large left sac is divided between a blind dome (fundus) rising above the cardia and a body (corpus) extending from the cardia to the ventral angle. On the basis of the terminal muscular thickenings, the more tubular right or pyloric part is divided between a more proximal pyloric antrum and a more distal pyloric canal (Fig. 3.35B). The margin separating the two surfaces is divided between greater and lesser curvatures, each of which runs between the cardiac and pyloric openings. The convex greater curvature attaches to the greater omentum, of which a part
(gastrosplenic ligament) connects the spleen with the stomach. The shorter, concave lesser curvature is connected with the liver by the lesser omentum. This curvature is marked by a sharp change in direction known as the angular notch (incisura).
The stomach wall is composed of layers corresponding to those of the esophagus and intestine.
The external peritoneum or serosa covers the entire organ, adhering to the underlying muscle, except along the curvatures, where it is reflected to continue into the omenta; its absence from the curvatures makes them the parts most likely to burst when the organ is excessively distended.The next coat is of smooth muscle and is arranged in three overlapping but incomplete layers. The external layer is more or less longitudinal, continues the outer muscle of the esophagus, and is concentrated along the curvatures, although it spreads more widely over the pyloric part. The middle layer is disposed in hoops, and those most proximal form a weak sphincter around the cardia; beyond this the pattern is interrupted by the projection of the fundus, but it is resumed at a lower level. It then continues to the pyloric canal, where the hoops are bunched together on the lesser curvature, forming a muscular knot (that in some species produces an obvious projection into the lumen) and fanning out on the greater curvature to sometimes constitute proximal and distal pyloric sphincters. The innermost layer is very incomplete but compensates for the deficiencies in the circular muscle; particularly stout fascicles arch above the cardia before continuing distally to each side of the lesser curvature, extending toward, but not beyond, the angular notch (Fig. 3.37).
The thin submucosa internal to the muscle is separated from the mucosa proper by a plexiform muscularis mucosae. It contains major arterial and venous plexuses and also a wealth of elastic fibers that help the muscularis mucosae throw the mucosa of the empty organ into the folds (rugae) that provide the characteristic surface relief (Fig. 3.37 and Fig. 3.38A). These folds are predominantly longitudinal in orientation, although individually tortuous; they are completely effaced only when the stomach is grossly distended.
The entire gastric mucosa has innumerable gastric pits. The pits, invisible to the naked eye, are depressions but account for the surface folding seen in histologic sections (Fig.
3.38B). The surface epithelium of columnar, mucus-secreting cells continues into the pits and even extends into the uppermost parts of the gastric glands that deliver their products into the depth of the pits. This epithelium is largely responsible for the protective coat that makes gastric mucosa slimy to the touch. The gastric glands are of three varieties, termed cardiac, proper gastric (fundic), and pyloric, although it must be stressed that in many species, including the dog, their distribution does not exactly coincide with the gross regions bearing the same names. The cardiac and pyloric glands produce additional mucus, whereas the proper gastric glands alone produce pepsin and hydrochloric acid, which are important parts of digestive juice. The enzyme is produced by the more numerous (chief) cells, the acid by the fewer parietal cells. There also are mucus-secreting cells. It is claimed that the proper gastric glandular region has a somewhat darker hue than the remainder of the mucosa.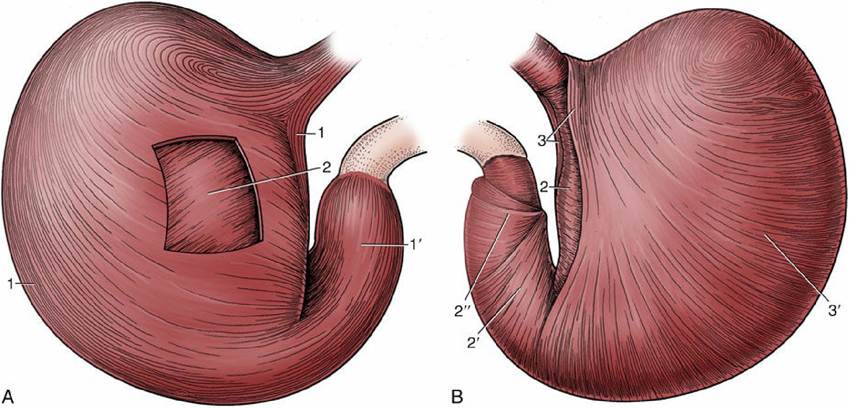
FIG. 3.37 The tunica muscularis of the canine stomach. (A) Parietal surface after removal of the serosa. (B) Stomach turned inside out with the mucosa removed. The tunica muscularis comprises outer
longitudinal, middle circular, and inner oblique layers. The longitudinal layer clothes the curvatures (1) and the pyloric part (1) but is thin over the body. The circular layer surrounds the body (2) and is especially prominent on the pyloric part (2), where it furnishes the pyloric sphincters (2"). The oblique layer (3) is thickest along the lesser curvature, where it forms two lips that fuse over the cardia (cardiac loop); it is thin where it lines the fundus and body (3).
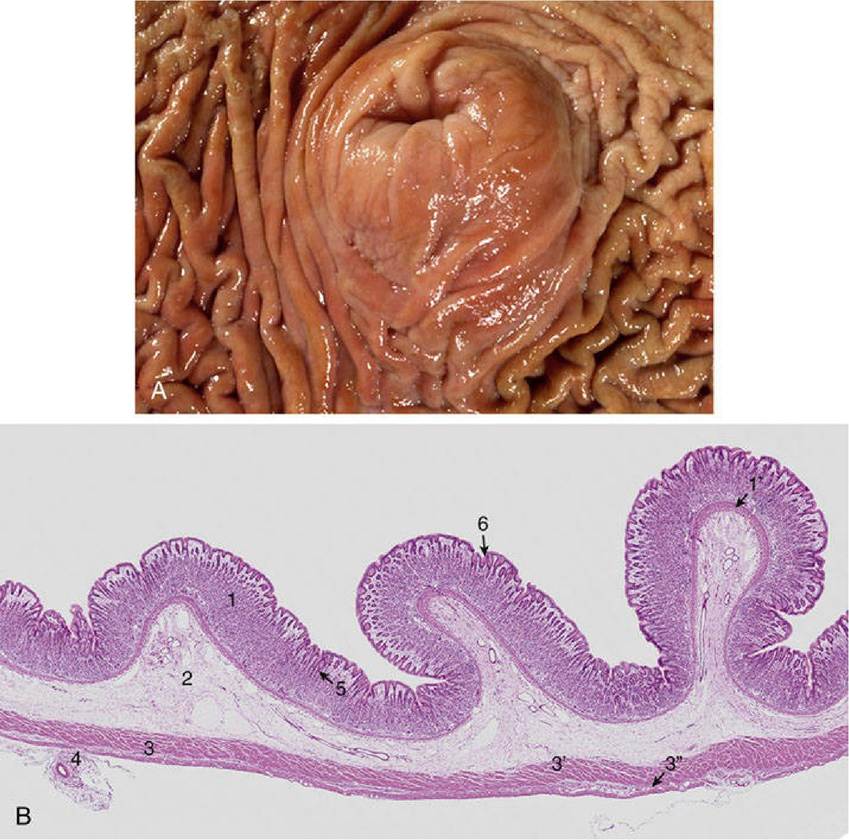
FIG. 3.38 (A) Protruding cardia surrounded by longitudinal folds or rugae. (B) Glandular region of stomach of cat.
Glandular region shows 3 rugae or folds. 1, tunica mucosa; 1,, lamina muscularis mucosae; 2, tunica submucosa; 3, tunica muscularis composed of inner oblique (3,) and outer longitudinal (3”) layers; 4, tunica serosa is thin with a blood vessel; 5, gastric glands located in lamina propria; 6, gastric pit (hematoxylin and eosin stain).The blood supply to the stomach comes from all three chief branches of the celiac artery and is particularly generous along the two curvatures (Fig. 3.39). The arteries anastomose quite freely externally and also within the stomach wall. For the most part, the arteries that penetrate the wall pass to the submucosa before branching to form an elaborate plexus from which both the muscular and the mucosal coats are fed. The mucosal branches supply unusually wide-bored capillaries below the epithelium and about the glands.
The veins are similarly arranged and ultimately combine to form trunks that join the portal vein. Numerous arteriovenous anastomoses provide a means of regulating mucosal blood supply, and much blood is diverted from the capillary bed of the fasting organ.
Lymph vessels are present in profusion, particularly in the submucosa. They lead to several gastric nodes, each charged with the drainage of a particular territory.
The stomach is innervated by parasympathetic fibers within the two vagal trunks and by sympathetic fibers that reach the organ with the arteries. The efferent fibers of both sets are accompanied by more numerous afferent fibers. Parasympathetic fibers of the vagus nerve synapse on ganglion cells in intramural plexuses within the submucosa and between the muscle coats and exert a high measure of control over gastric motility. In the proximal stomach, vagal activity suppresses muscular contraction and leads to adaptive relaxation, whereas in the distal stomach, vagal stimulation causes intense peristaltic activity. Vagal stimulation of distal antral motility is mediated by acetylcholine, but the identity of the inhibitory mediator is not well established and it may be vasoactive intestinal peptide.
The intramural plexuses are involved in the local reflexes in which the stomach wall reacts to direct stimulation. Sympathetic and parasympathetic fibers also innervate the surface epithelium and glands, but only parasympathetic fibers end on the intragastric endocrine cells.* Division of the vagal nerves, either the main trunks or selected branches, reduces gastric activity and secretion.* There are several varieties of these cells, the most important of which is the gastrin-secreting cells scattered singly within the epithelium of gastric glands, especially those of the antral region. The release of gastrin is stimulated by the vagal nerves and also, more directly, by distention of the stomach by a meal. Gastrin is passed into the portal circulation, returning within the arterial blood to promote increased activity, both glandular and muscular, of the stomach wall.
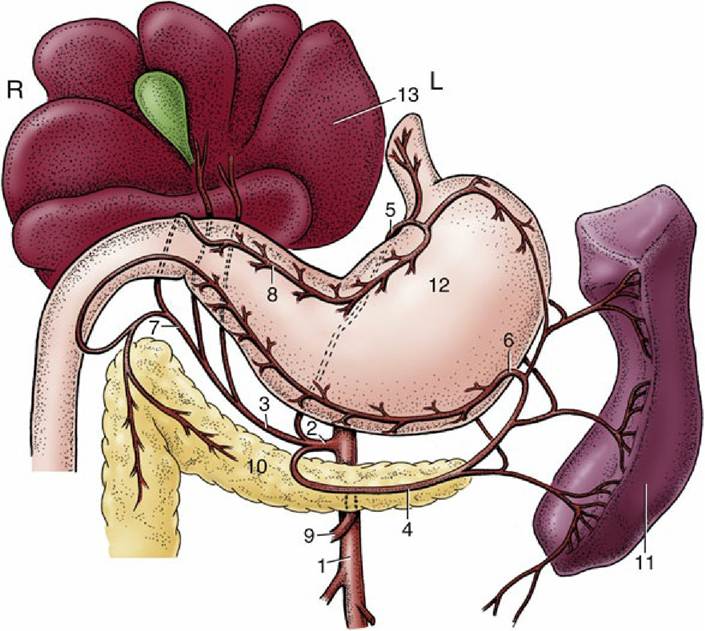
FIG. 3.39 Distribution of the celiac artery of the dog (ventral view). 1, Aorta; 2, celiac artery; 3, hepatic artery; 4, splenic artery; 5, left gastric artery; 6, left gastroepiploic artery; 7, gastroduodenal artery; 8, right gastric artery; 9, cranial mesenteric artery; 10, pancreas; l1, spleen; l2, stomach; 13, liver; L, left; R, right.
The empty stomach is small and contracted toward the fixed point of the esophageal entrance such that it is entirely within the rib cage and fails to reach the abdominal floor. The wall is generally inert except for occasional weak peristaltic contractions, and little secretion from the glands occurs. Any residual peristaltic activity ceases as soon as food is offered (or anticipated). Secretion increases as a reflex response to the taste of food or the effort of mastication. When food does arrive, it first collects in layers (because as yet no mixing movements are present) largely in the body, to expand it in all directions but principally ventrally and caudally. A motor response is delayed and slowly builds to a peak. Peristaltic contractions commence near the cardia and course distally, accelerating and becoming more vigorous when they reach the muscular pyloric antrum. The terminal segment contracts en masse to move the ingesta into the duodenum while the peristaltic wave is still some distance from the pylorus. Radiographic studies suggest that the pylorus is open for about one third of the time. It is probable that emptying is dependent more on intermittent increase in the intragastric pressure than on the regular peristaltic activity.
The effects of feeding on topography and relations of stomach with other organs are considerable, especially in animals kept under regimens that allow them to feed seldom but to repletion. The fully distended stomach may extend almost to the umbilicus—or even beyond this in the puppy—pushing the intestinal mass dorsally and caudally. The liver is pushed to the right while the spleen, tethered to the left part of the greater curvature, follows the expansion of that side of the stomach.