THE STRUCTURAL ELEMENTS
An appropriate environmental change provides a stimulus that is recognized by a receptor organ; the reaction or response that may be provoked in answer to the stimulus is performed by an effector organ (Figure 8-1).
In multicellular organisms the receptor and effector organs are separate and are connected by a chain of neurons, highly specialized cells in which the general cytoplasmic properties of excitability and conductivity are developed to extreme degrees. Whatever the stimulus, the receptor neuron translates it into an electrical potential, and the message is transmitted in this coded form. The impulse travels the length of the neuron before transmission to the next cell in the chain; this may be another neuron or interneuron, but ultimately, at the end of the chain, the motor neuron will end on an effector muscle or gland cell. Neurons thus provide the basic units from which the nervous system is constructed.The typical neuron is an elongated cell that consists of a cell body containing the nucleus (therefore known as the perikaryon) and various processes (Figure 8-2). The processes, which vary considerably in number, length, and form, are of two varieties obviously and usefully distinguished by the direction in which they transmit impulses. One variety, the dendrite, is usually multiple and transmits impulses toward the perikaryon; the other, the axon, is always single at its origin (although it may divide at some distance from the perikaryon) and conveys impulses away. The nerve cell is thus clearly polarized. The arrangement of the processes permits a simple morphological classification of neurons. Most are multipolar and possess a number (often a very large number) of branching dendrites that join the perikaryon at scattered points (see Figure 8-2). In a second type, bipolar, the dendrites join in a common trunk before reaching the perikaryon at a site remote from the origin of the axon.
In the third type, unipolar, the dendrite tree and axon first combine in a single extension of the perikaryon that later branches; such neurons are also described as pseudounipolar because they initially develop as bipolar cells. Dendrites and axons are superficially alike, and both are commonly described as nerve fibers. As a general rule to which there are many exceptions, dendrites are relatively short and axons relatively long.The different varieties of a neuron have specific distributions that are related to their particular functions. Clearly, a much-branched dendritic tree enables a neuron to receive impulses from many sources. Conversely, a much-branched axon makes connection with and stimulates many cells. The first arrangement allows a convergence of impulses from various origins; the second provides for a divergence or diffusion of a message.
Interneuronal connections are known as synapses, a term usually broadened to also include neuromuscular connections. An axon may establish synaptic connections with the bodies, dendrites, or axons of other neurons, which are varieties of synapses distinguished as axosomatic, axodendritic, and axoaxonic. Most neurons establish many synapses: some have many thousands of synaptic sites, though not in relation to so many other cells. Synapses have a variable but always complicated morphology, but only an elementary description is required here. The participating cells are neither continuous nor in direct contact but are always separated by a very narrow gap. A nerve impulse (action potential) arriving at the presynaptic part of an axon does not jump from cell to cell; instead, it prompts the release of a specific chemical transmitter substance that diffuses across the gap. When this substance arrives at the postsynaptic plasma membrane (of the following cell), it produces one of two effects: it either depolarizes
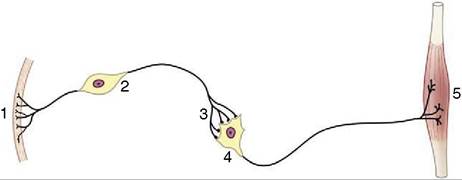
Figure 8-1 A simplified receptor-effector system.
1, Skin receptor; 2, afferent neuron; 3, synapse; 4, efferent neuron; 5, striated muscle (effector organ).the membrane, initiating a fresh impulse, which is then propagated the length of the postsynaptic cell, or it hyperpolarizes the membrane, producing a blocking or inhibitory effect. The existence of both excitatory and inhibitory synapses, sometimes on the same cell, provides a means for a great diversity of response. Many transmitter substances are known; the most common include acetylcholine, glutamate (excitatory), GABA (inhibitory) noradrenaline, serotonin, and many neuropeptides.
Neurons are supported by other specialized cells. The supporting tissue of the brain and spinal cord is known as neuroglia and comprises several cell types that we shall not distinguish. Neuroglial cells not only support the neurons but also assist in their nutrition and neurotransmission; additionally, neuroglial cells provide nerve fibers within the brain and spinal cord with cytoplasmic products that insulate them from their surroundings and prevent leakage of the impulses they convey. The insulating material, myelin, incidentally imparts a white color to nerve fibers seen en masse. Nerve fibers within peripheral trunks (outside the brain and spinal cord) receive similar insulation—of very variable thickness— from another type of supporting cell, the Schwann cell (neurolemmocytus; Figure 8-3). Peripheral nerve trunks are further protected, supported, and subdivided by
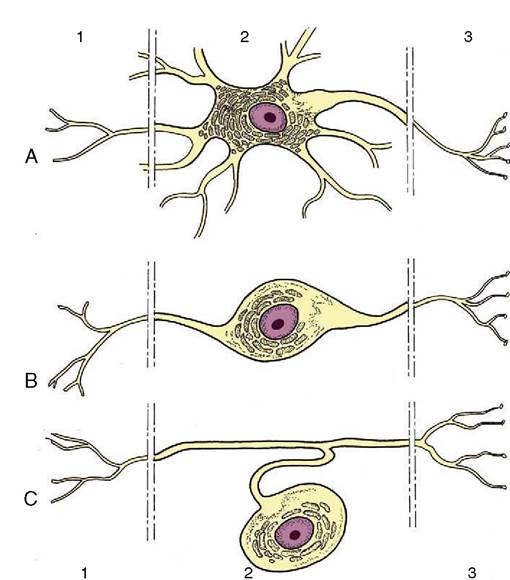
Figure 8-2 Multipolar (A), bipolar (B), and pseudounipolar (C) neurons. 1, Receptor side (dendrites); 2, cell body (perikaryon); 3, effector side (axon).
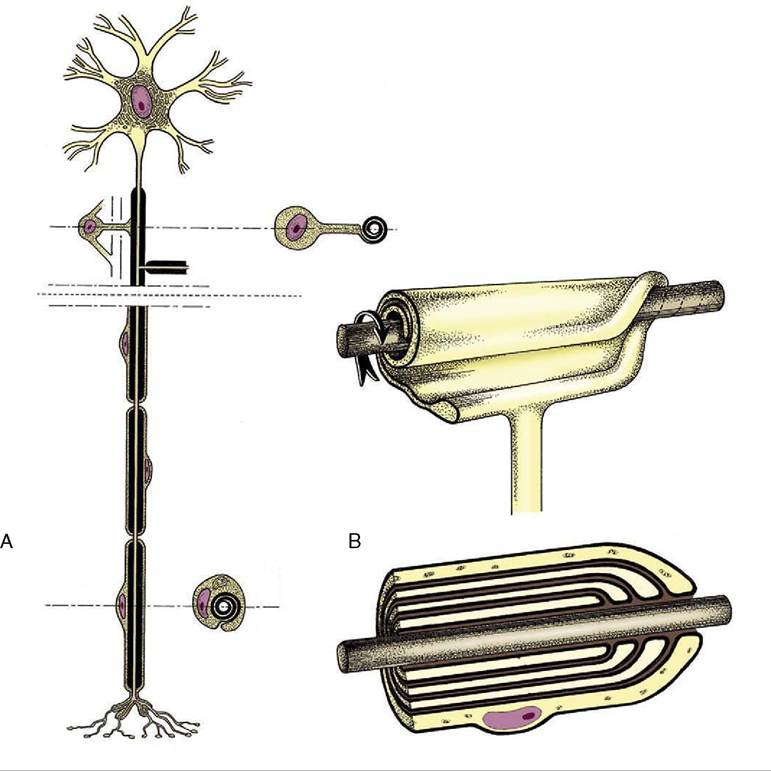
Figure 8-3 A, Neuron with its axon enwrapped within a cytoplasmic sheath supplied by a series of Schwann cells. B, The cell membrane of the Schwann cell is rolled around the axon.
The investment may consist of several plasmalemma layers forming a thick myelin sheath.connective tissue sheaths and septa, but the brain and cord, although included within a series of connective tissue investments (meninges), are not penetrated by connective tissue in this way.
Groups of perikarya are distinguished by their darker color, especially when set off by the whiteness of adjacent fiber bundles; this permits the ready distinction of the “gray” (in fact beige in the fresh specimen) and white substance of the brain and cord. Isolated neuronal aggregations within the brain are generally known as nuclei; many are too small to be distinguished by the naked eye.
Fiber bundles of common origin, destination, and function tend to be aggregated within the brain and cord into fasciculi or tracts, although the limits of these are not normally evident and can be made so only by experimental means. Most such tracts are named by the combination of their origin, employed as prefix, with their destination, employed as suffix; the significance of such names as the spinocerebellar and cerebellospinal tracts is thus revealed directly.
Neuronal aggregations on peripheral nerves may form visible swellings; they may also be distinguished by their color and texture, which are darker and firmer than those of the related nerve trunks. They are universally known as ganglia.