STIMULUS-RESPONSE APPARATUS
Having established these fundamental points, we may now return to consider the stimulus-response apparatus. In the simplest form found in mammals, this apparatus comprises five elements arranged in series: a receptor region adapted to respond to a stimulus of a particular
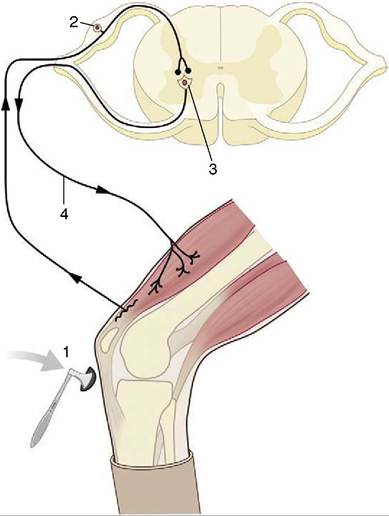
Figure 8-4 The monosynaptic patellar reflex.
The stretch stimulus on the tendon (1) travels via the afferent neuron (2) to the spinal cord. The impulse is then transmitted to the efferent neuron (3), which stimulates the quadriceps muscle (4).modality (sound, touch, and so forth); an afferent neuron that conveys an impulse centrally, toward the brain or cord; a synapse; the remainder of the efferent neuron that conveys an impulse from the center to the periphery; and an effector, which may be a muscle, gland, or neurosecretory cell (see Figure 8-1). This sequence constitutes a primary, elementary, or monosynaptic reflex arc.
The monosynaptic reflex arc is actually a most uncommon arrangement, although it does provide the basis of one familiar example, the patellar or knee-jerk reflex (Figure 8-4). This is a stretch (myotactic) reflex that, as many readers will know, can be elicited by an appropriate tap on the patellar ligament, which is the functional continuation of the quadriceps femoris muscle. The tap stretches the muscle and thus stimulates muscle spindles and other receptors within its belly and tendon; an impulse travels along afferent fibers within the femoral nerve to reach the spinal cord, where it is projected on efferent (lower motor) neurons. The axons of these neurons return within the femoral nerve, and the impulse is then projected on the constituent fibers of the muscle,
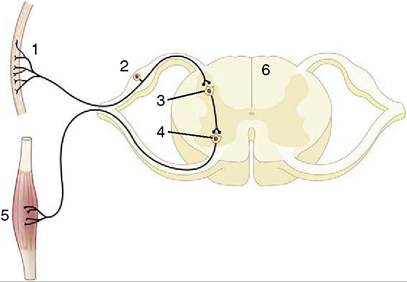
Figure 8-5 Schematic representation of a reflex chain in which an interneuron is interposed.
1, Skin receptor; 2, afferent neuron; 3, synapse at interneuron; 4, synapse at efferent neuron; 5, muscle; 6, spinal cord.stimulating their contraction to effect the abrupt extension of the joint.
In most reflexes one or more additional neurons are interposed in the chain between the afferent and efferent neurons (Figure 8-5). These are conveniently known as interneurons, although several synonyms exist. The system may still be described as simple if only an unbranched neuronal chain is involved. However, most reflexes involve more complicated circuitry in which additional neurons are stimulated (or inhibited). Collateral branching enables the exercise of a more refined control and possibly the intrusion of the activity on consciousness.
A good example of an integrated response is given by the limb of a standing animal subjected to a prick or other noxious stimulus. The limb is withdrawn by the coordinated action of the flexor muscles of several joints; these movements are facilitated by the relaxation of the previously active and antagonistic extensor muscles. The branching pathways involved in securing this response extend through several segments of the cord to reach and excite, or inhibit, the efferent neurons that supply the various muscles. At the same time, the animal has to adjust to the removal of one of its supporting props by redistributing its weight over the other limbs; the pathways necessary for this wider adjustment extend through considerable stretches of the cord, some of which cross to the contralateral side (Figure 8-6).
Coordination of the changes so that balance may be maintained involves higher centers within the brain, to which the message must ascend, in addition to integration within the cord. The process is unlikely to go unnoticed; the cortex is involved, and the animal assesses the situation and considers whether a more general response, such as flight or retaliation against the aggressor, would be appropriate. This considered response is a far cry
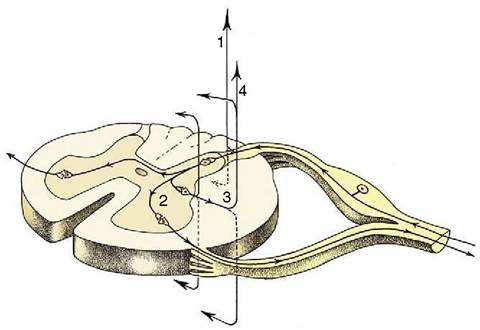
Figure 8-6 The course of fibers within the spinal cord.
Some afferent fibers in the dorsal funiculus travel directly toward the brain (1); others end on interneurons in the dorsal horn. From here impulses can be transmitted directly to efferent neurons (2) or to other interneurons that transmit impulses caudally or cranially within the spinal cord (3), some extending to the brain (4).from the simple, monosynaptic, and monosegmental response of the knee jerk and involves integrative apparatuses of various degrees of complexity, spread through the cord and brain, and drawing on those higher centers that are concerned with memory and judgment.
THE SUBDIVISIONS OF THE
NERVOUS SYSTEM quently termed sensory; the impulses travel from the periphery toward the brain or spinal cord. Within the cord they are more often described as ascending; the impulses travel from “lower” (more caudal) toward “higher” (more cranial) parts. Efferent pathways usually conduct impulses from “higher” to “lower” levels within the brain and cord and from these to the periphery; the alternative names to describe these systems are descending and motor. The equivalence of certain of these terms does not withstand close scrutiny, particularly when applied to integrative systems within the spinal cord; many descending fiber bundles are not motor, and many ascending bundles are not sensory.
The nature of the information that is conveyed, as well as the nature of activities that are directed, permits the further distinction of somatic and visceral nervous systems. The somatic system is concerned with those functions, like locomotion, that determine the relationship of the organism to the outside world. The visceral system is concerned with functions that relate to the internal environment: the regulation of the vascular system and heart rate, the control of glandular activity and digestive processes, and so forth. As a general but not invariable rule, there is a greater awareness and greater voluntary control of somatic than visceral functions; of course they work in close collaboration.
A more elaborate classification is possible. Afferent systems are initially divisible into somatic and visceral divisions, and these in turn are divisible into general and special subdivisions.
Somatic afferent pathways originate in receptors within the skin and deeper somatic tissues of the body wall and limbs. The pathways that arise from skin receptors are concerned with the exteroceptive sensations, such as touch, temperature, and pain, that respond to stimuli delivered from outside the organism. Receptors within the deeper tissues include the additional proprioceptive category concerned with such “deep” sensations as those that inform on the present angulation of the joints and tension within muscles and tendons and on changes in these conditions. Somatic afferent fibers are carried by all spinal nerves and by the fifth cranial (trigeminal) nerve (see Table 8-2, p. 286).
Special somatic afferent pathways have a more restricted origin within certain special sense organs: the retina of the eye and the cochlear and vestibular components of the inner ear, which are concerned with vision, hearing, and balance, respectively. The fibers concerned with vision and hearing are exteroceptive, those concerned with balance proprioceptive. Special somatic afferent fibers are thus found only within two cranial nerves, the optic and vestibulocochlear nerves.
Visceral afferent pathways originate in the (entero- ceptive) receptors of vessels and glands and the viscera of the head and trunk that mostly respond to stretch and chemical stimuli. The fibers of this division are found in the cranial nerves III, V, VII, IX, and X, certain sympathetic and parasympathetic nerves, and all spinal nerves.
Special visceral afferent pathways arise from the special sense organs of smell and taste. Fibers conveying olfactory information are confined to the olfactory nerve; those conveying gustatory (taste) information are confined to a small group of cranial nerves.
Efferent systems are divided more simply.
Somatic efferent pathways lead to striated muscles of somitic and branchiomeric origin.
Visceral efferent pathways lead to the smooth muscle of the viscera and vessels, to heart muscle, and to glands. Most of these organs receive a double innervation through the sympathetic and parasympathetic divisions of the autonomic nervous system (p. 327), which are often described as antagonistic, although “balancing” might better suggest their cooperative role. Visceral efferent fibers of the sympathetic division leave the central nervous system via the spinal nerves in the thoracolumbar regions of the cord; those of the parasympathetic division are limited to a small group of cranial nerves and to the sacral contingent of spinal nerves. However, many visceral efferent fibers later join other nerves so that they finally obtain a very widespread peripheral distribution.
Somatotopy
The fibers and cell bodies within many tracts and relaying nuclei and within areas of the cerebral and cerebellar cortices on which these may project preserve very orderly point-to-point arrangements that reflect the topography of the parts of the body from which afferent impulses arise or to which efferent impulses are delivered. These do not always, or even usually, reproduce the true proportions but represent the parts of the body in relation to the densities of their innervation. The representations take the form of grotesque caricatures, sometimes known as homunculi—although animalcula would better fit veterinary anatomy—in which very sensitive parts, such as the lips and muzzle of the horse, or those capable of very refined and accurate movements, such as the fingers of a human or the prehensile tail of a monkey, are of exaggerated size. The concept of somatotopy is of great importance in the consideration of the significance of pathological lesions, in the conduct of neurosurgery, and in experimental stimulation.
*This term identifies structures originating in the serial pharyngeal (branchial) arches.