The Ventilatory Response to Carbon Dioxide Is Mediated Through a Medullary Chemoreceptor
By probing with CO2 or acetazolamide (an inhibitor of carbonic anhydrase), chemosensitive tissue has been located along the ventral medulla, near the nucleus tractus solitarius, in the region of the ventral respiratory group of neurons, and also in regions of the brain not known to have a function in control of breathing.
These chemosensitive neurons (the central chemoreceptor) apparently respond to changes in the pH of the interstitial tissue fluid in which they are bathed. A decrease in pH increases ventilation, and an increase in pH decreases ventilation. The brain interstitial fluid, which bathes the central chemosensitive neurons, communicates directly with cerebrospinal fluid (CSF). Because of this, changes in ventilation can be induced by changes in the composition of arterial blood and experimentally by changes in the pH of CSF (Figure 49-5).The central Chemosensilive neurons are separated from blood by the blood-brain barrier, which is freely permeable for CO2 but less permeable for hydrogen ions (H+) and bicarbonate (HCO3"). An increase in blood Pco2 causes a rapid increase in Pco2 in the brain interstitial fluid. Carbonic acid (H2CO3) forms and dissociates into H4 and HCO3" because the interstitial fluid is poorly buffered, the pH around the chemosensitive neurons decreases, and this stimulates ventilation. Similarly, infusing H‘ into the CSF decreases the interstitial fluid pH and increases ventilation.
An acute increase in the blood H+ concentration is not reflected immediately by a decrease in the pH of either the interstitial fluid or CSF, because the blood-brain barrier is relatively impermeable for H‘. Therefore, acute increases in blood H ’ concentration are detected by the peripheral chemoreceptors. However, changes in the brain interstitial fluid pH may follow those in the blood within K) to 40 minutes.
The composition of CSF and thus brain interstitial fluid has a major effect on the response of the central chemoreceptor. If the HCOf concentration of the CSF decreases, as occurs in metabolic acidosis, the buffering capacity of the CSF is reduced. An increase in Pco2 then causes a greater decrease in the pH of the CSF than would occur in the presence of a normal buffering capacity, and thus the ventilatory response to CO2 is more vigorous than normal (Figure 49-6). Conversely, in metabolic alkalosis, the HCO3- concentration in CSF increases, and the response to CO2 is depressed. The CSF composition is regulated by the active transport of ions at the choroid plexus. In metabolic acidosis and alkalosis, changes in the HCOf concentration in CSF tend to follow changes in the blood HCO3" concentration, but with a phase lag of several hours.
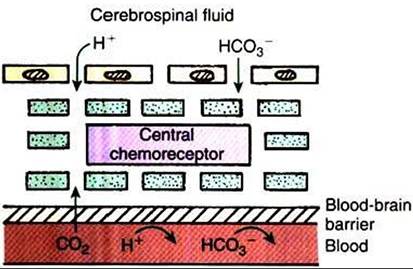
FIGURE 49-5 ■ Diagrammatic representation of the central chemoreceptor separated from blood by the blood-brain barrier, which allows passage of carbon dioxide (CO2) but is less permeable for hydrogen ion (H*) and bicarbonate (HC03^).The central chemoreceptor is bathed by brain interstitial fluid, which is in communication with cerebrospinal fluid.
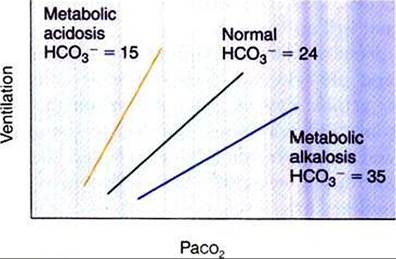
FIGURE 49-6 Effect of arterial carbon dioxide tension (Paco2) on ventilation in a normal animal and in animals with metabolic acidosis and alkalosis. A decrease in the bicarbonate (HCO3) level (mEq∕L) increases the ventilatory response to carbon dioxide.