The Ventral Rami
The larger ventral rami supply the hypaxial muscles, including those of the limbs (excepting the thoracic girdle muscles supplied by the eleventh cranial nerve and the rhomboideus supplied in some species by dorsal rami) and the remaining skin of the neck, trunk, and limbs.
Except in the thoracic region, where a more precise segmental distribution is retained, the ventral rami are also joined with their cranial and caudal neighbors by connecting branches. These connections are greatly exaggerated at the levels of origin of the nerves to the forelimb and hindlimb, where they constitute the brachial and lumbosacral plexuses, respectively.The Cervical Ventral Rami
The cutaneous distribution of the first two cervical ventral rami extends rostrally to the external ear and the masseteric and throat regions. The more caudal cervical ventral rami, in addition to providing sensory innervation to dermatomes of the neck region, also provide motor axons to the phrenic nerve and to the brachial plexus, discussed later.
In domestic species the phrenic nerve, the nerve supplying the diaphragm, is generally formed by the fifth, sixth, and seventh cervical nerves. Axons that contribute to the phrenic nerve leave the large ventral rami to run ventrally over the scalenus muscle to join in a trunk (see Fig. 1.38) that winds below the muscle to enter the mediastinum between the two first ribs. The phrenic nerve runs caudally within the mediastinum, crossing the lateral surface of the pericardium, to reach the diaphragm; the right nerve utilizes the plica venae cavae in the last part of its course (see Fig. 13.14/12 and Fig 13.15/6). The phrenic nerves ramify within the diaphragm to which they are the sole motor innervation; their sensory fibers are supplemented by others channeled through intercostal nerves. It is worth emphasizing that the diaphragmatic muscle is skeletal muscle and phrenic nerves contain somatic efferent fibers; it must not be inferred from the normally involuntary nature of breathing that phrenic nerves contain visceral efferent—in other words, autonomic—fibers.
Experiments in some species have shown that bilateral section of the phrenic nerves has little effect, although respiratory distress may become evident when the animal is severely stressed.The Brachial Plexus
The brachial plexus is a network of nerves that supplies sensory and motor innervation to almost all structures of the forelimb—except for the trapezius, omotransversarius, brachiocephalicus, and rhomboideus — and the skin over the upper shoulder region.
The plexus is usually formed by contributions from the last three cervical and first two thoracic nerves; the fifth cervical nerve sometimes participates, and the contribution of the second thoracic nerve is then reduced or lacking. The plexus reaches the axilla by passing between the parts of the scalenus and quickly splits into peripheral nerves that diverge toward their separate destinations (Fig. 8.74). Several of these nerves have very restricted local distributions, and bare mention of their names and destinations is all that is required; they include the long thoracic nerve (Fig. 8.74/9) to the serratus ventralis, the thoracodorsal nerve (Fig. 8.74/9') to the latissimus dorsi, the cranial and caudal pectoral nerves (Fig. 8.74/3 and 9") to the pectoral muscles (including the subclavius), the subscapular nerve (Fig. 8.74/2) to the subscapularis, and the lateral thoracic nerve (Fig. 8.74/9") to the cutaneous trunci and to skin over the ventral part of the thorax and abdomen. The other nerves, described here, require fuller description. There are some interspecies differences, but these are rarely of importance except in the manus.
The suprascapular nerve (Fig. 8.74/1) leaves the cranial part of the brachial plexus (C6-C7). It passes between the supraspinatus and subscapularis to reach the cranial margin of the neck of the scapula, around which it winds to the lateral aspect of the bone, where it is distributed to supply the supraspinatus and infraspinatus muscles. Like other nerves directly apposed to bone, it is vulnerable to injury; in this case it is usually stretched against the scapula when the limb is overabducted or violently retracted.
The resulting paralysis of the lateral shoulder muscles does not affect the standing posture but may result in an obvious lateral movement of the shoulder joint ("shoulder slip") during the stride. The condition occurs most frequently in horses, in which it is also known as "sweeny"; it manifests itself after a time by obvious wasting of the muscles beside the scapular spine.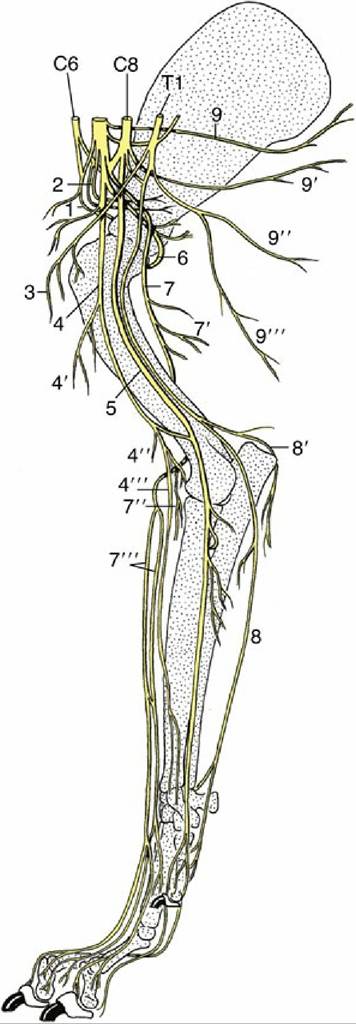
FIG. 8.74 The nerves of the right forelimb of the dog; medial view. 1, Suprascapular nerve (n.); 2, subscapular nerves (nn.); 3, cranial pectoral nn.; 4, musculocutaneous n.; 4', proximal muscular branch; 4", distal muscular branch; 4'", medial cutaneous antebrachial n.; 5, median n.; 6, axillary n.; 7, radial n.; 7', muscular branches to triceps; 7", muscular branches to extensors; 7''', cranial cutaneous antebrachial n.; 8, ulnar n.; 8 ', caudal cutaneous antebrachial n.; 9, long thoracic n.; 9 ', thoracodorsal n.; 9", lateral thoracic n.; 9''', caudal pectoral n.; C6 and C8, sixth and eighth cervical nn.; 71, first thoracic n.
The musculocutaneous nerve (Fig. 8.74/4) is also of cervical origin (C7-C8). After a short course within the axilla, the nerve provides a proximal branch (Fig. 8.74/4'), which supplies and terminates in the coracobrachialis and biceps in the upper part of the arm. In the dog, the main part of the nerve continues into the distal third of the arm, where a communicating branch passes distocaudally to the median nerve. The remaining trunk of the musculocutaneous nerve passes under the terminal part of the biceps brachii, where it forms a distal branch (Fig. 8.74/4'), which supplies the brachialis, and the medial cutaneous nerve of the forearm (Fig. 8.74/4"), which crosses the flexor aspect of the elbow before ramifying in skin.
In ungulates, the musculocutaneous nerve loops around the axillary artery to join the median nerve; further distally, the musculocutaneous fibers again separate from the median nerve in the upper and lower parts of the arm, where they form the proximal and distal muscular branches of the musculocutaneous nerve.
In the horse alone, the cutaneous branch extends beyond the carpus to the fetlock.Damage to the main musculocutaneous trunk is an unlikely injury; it would paralyze the main flexors of the elbow, although compensation would probably be found from activity of the carpal and digital extensors.
The axillary nerve (C8) (Fig. 8.74/6) passes behind the shoulder joint to reach the lateral aspect of the limb. En route it supplies the teres major, teres minor, capsularis, and deltoideus—the true flexors of the shoulder joint. It also supplies small branches to the distal part of the brachiocephalicus, which, it will be recalled, is of deltoid origin. A cutaneous branch supplies skin over the cranial aspect of the arm and forearm.
The three remaining branches of the plexus have the most complicated courses and the most extensive distributions. The radial nerve (Fig. 8.74/7) arises from the last two cervical and first thoracic nerves (C7-T1). It first runs distally within the arm, caudal to the brachial artery, before diving between the long and medial heads of the triceps to follow the spiral groove of the humerus, which leads it to the craniolateral aspect of the limb. The nerve supplies branches to the various heads of this muscle (Fig. 8.74/7') and to tensor fasciae antebrachii and anconeus. In the lower part of the arm, the radial nerve supplies a further set of branches (Fig. 8.74/7") to all carpal and digital extensor muscles, including the anomalous ulnaris lateralis. A cutaneous branch or branches (Fig. 8.74/7) descends over the craniolateral aspect of the forearm and carpus to reach the dorsal surface of the digits. In the horse, this cutaneous contribution ends at the level of the carpus, and more distal innervation is assumed by the musculocutaneous nerve.
Damage to the radial nerve can have three obvious consequences: paralysis of the elbow extensors, paralysis of the carpal and digital extensors, and anesthesia of the denervated skin regions. The combination of all three disabilities points to injury proximal to the middle of the arm, the combination of the second and third points to injury in the distal part of the arm, and a purely sensory deficit suggests injury beyond the origin of the distal motor branches.
Injury in the arm is quite common because in places only a thin layer of muscle separates the radial nerve from the humerus, and the nerve may be involved in fracture or tumor of this bone. Extensive damage to the radial nerve proximal to the origin of the branches to the triceps is serious because it prevents fixation of the elbow, prohibiting the limb from bearing weight; the foot is dragged with its dorsal surface on the ground. More distal lesions are less serious because the elbow can be fixed, and most animals learn to compensate for paralysis of the forearm muscles by flicking the limb forward and planting the foot quickly to prevent toe drag.The median nerve (Fig. 8.74/5) comes mainly from the last cervical and first thoracic nerves (C8- T1). It runs down the medial surface of the arm caudal to the main artery and enters the forearm muscle over the medial collateral ligament of the elbow joint, where it provides motor innervation to many of the carpal flexors. It inclines caudally, passes under the flexor carpi radialis, and maintains this protected situation until it reaches the carpus. It divides in the distal part of the forearm, or within the carpal canal, into two or more divisions that descend through the carpal canal to supply most structures of the palmar part of the foot. The median nerve supplies most of the flexor muscles of the carpus and digit in a pattern that overlaps (but does not quite coincide) with the distribution of the ulnar. Therefore, damage confined to the median nerve is not usually manifested through any abnormality of posture or gait.
The ulnar nerve (Fig. 8.74/8) leaves the caudal part of the plexus (C8-T2). It runs down the arm adjacent to and possibly conjoined with the median nerve before deviating in the direction of the olecranon to cross the caudal aspect of the elbow joint. Within the arm it detaches the caudal cutaneous antebrachial nerve. The main trunk is severely depleted by detachment of the branches to the carpal and digital flexor muscles in the upper part of the forearm, and the narrow continuation runs down the caudal aspect of the forearm.
It finally divides a short distance above the accessory carpal bone. The dorsal branch of this division emerges between the tendons of the ulnar carpal flexor and ulnaris lateralis and descends over the lateral face of the accessory bone to supply the skin on the lateral aspect of the forefoot. The palmar branch continues through the carpal canal and later supplies the interosseous and other small muscles of the foot. It also supplies sensory branches to skin and deeper structures. The distribution within the foot is in close collaboration with the median nerve, partly through combined trunks. The innervation of the forefoot, a topic of considerable practical importance in horses, is later considered separately.Damage confined to the ulnar nerve is unlikely to impair locomotion; the sensory deficits show considerable interspecies variation.
The Thoracic Ventral Rami
The thoracic ventral rami show a more strictly segmental distribution than is found in other regions. The first two thoracic rami contribute to the brachial plexus, but generally the thoracic ventral rami provide the intercostal nerves that run ventrally within the intercostal spaces, either directly below the pleura or between the two intercostal muscle layers; the relation varies according to location and species. Apart from supplying the intercostal muscles, the intercostal nerves detach lateral cutaneous branches that supply a band of skin over the lateral aspect of the chest wall and ventral cutaneous branches that supply the skin of the ventral chest wall; the more caudal thoracic rami also supply sensory innervation to the abdominal floor. In the sow, bitch, and cat, the lateral cutaneous branches detach branches to supply sensory innervation to the thoracic mammary glands.
The last thoracic ventral branch (costoabdominal nerve) is slightly different in its course and distribution because it runs behind the last rib. It joins with the lumbar ventral branches to supply the flank.
The Lumbar Ventral Rami
The lumbar and sacral ventral rami form a continuous plexus, best developed where the last three or four lumbar and first two sacral nerves form the lumbosacral plexus that supplies the hindlimb. The more cranial lumbar ventral rami have a considerable importance in cattle because they are frequently blocked for abdominal surgery. They are given individual names; in species (including cattle) in which there are six lumbar nerves, the first ventral ramus is known as the iliohypogastric, the second is known as the ilioinguinal, and the third and fourth combine to form the genitofemoral nerve. In species with seven lumbar nerves the first two ventral rami are distinguished as the cranial and caudal iliohypogastric; the third supplies the ilioinguinal and also makes a contribution to the genitofemoral nerve. The genitofemoral nerve divides into a femoral branch that supplies the skin over the medial aspect of the thigh and a genital branch that supplies the spermatic fasciae, the scrotum, and the prepuce.
It is important to note that the ventral rami travel in a caudoventral direction rather than strictly ventral—this feature is most obvious in the lumbar rami but is apparent with the caudalmost thoracic rami, the intercostal nerves. Thus, the locations of their corresponding dermatomes and the locations where these nerves can most easily be accessed for injection of local anesthetic solution are both considerably more caudal than would naturally be supposed (see Fig. 28.2). The lumbar nerves pass through the transversus close to the tip of the transverse processes and then run deep to the internal oblique toward the abdominal floor (see Fig. 1.37). In addition to supplying the flank and rectus muscles, the lumbar nerves detach lateral and ventral cutaneous branches; the former appear subcutaneously at increasingly dorsal levels as the series is followed caudally.
The Lumbosacral Plexus
The lumbosacral plexus that gives origin to the nerves of the hindlimb (with the minor exceptions of those to certain proximal skin areas) is an enhancement of the continuous plexus described previously. It usually begins at the ventral ramus of the fourth lumbar nerve and ends with that of the second sacral nerve (L4-S2); it thus has an additional root in species possessing seven lumbar nerves (Fig. 8.75).
The femoral nerve (Fig. 8.75/1) arises from the cranial part (L4-L6) of the plexus and pursues a course through the psoas muscles to reach the gap between the dorsocaudal corner of the flank and the iliopsoas muscle. It is accompanied by the external iliac artery and vein, and on entering the thigh it runs in a protected position between the sartorius and pectineus. It soon detaches the saphenous nerve, and after a very short further course it dives between the rectus femoris and vastus medialis to branch within and innervate the quadriceps mass (Fig. 8.75/1'). Severe damage to this nerve, though relatively infrequent, has serious consequences because paralysis of the quadriceps prevents fixation of the stifle joint, rendering the whole limb incapable of supporting weight. No compensation for this defect is possible.
The saphenous nerve (Fig. 8.75/1") innervates the sartorius before continuing to supply skin over the medial aspect of the limb from the stifle to the metatarsus.
The obturator nerve (Fig. 8.75/2) has broadly the same origin (L4-L6) as the femoral nerve. It follows the medial aspect of the shaft of the ilium to reach the obturator foramen, through which it passes to the adductor muscles of the thigh; this group of muscles comprises the gracilis, pectineus, adductor, and obturator externus — and obturator internus in ruminants and the pig.* The close relationship of the obturator nerve to the pelvis is potentially dangerous, because it exposes the nerve to the risk of laceration in fractures and to the risk of compression during calving and foaling. The risk is less in species in which the young are small relative to the pelvic cavity. The effects of injury vary depending on the location of the damage along the extent of the nerve, but consequences of injury are greater in heavier animals and are exaggerated by a requirement to walk on smooth ground, when the limb tends to slip sideways.
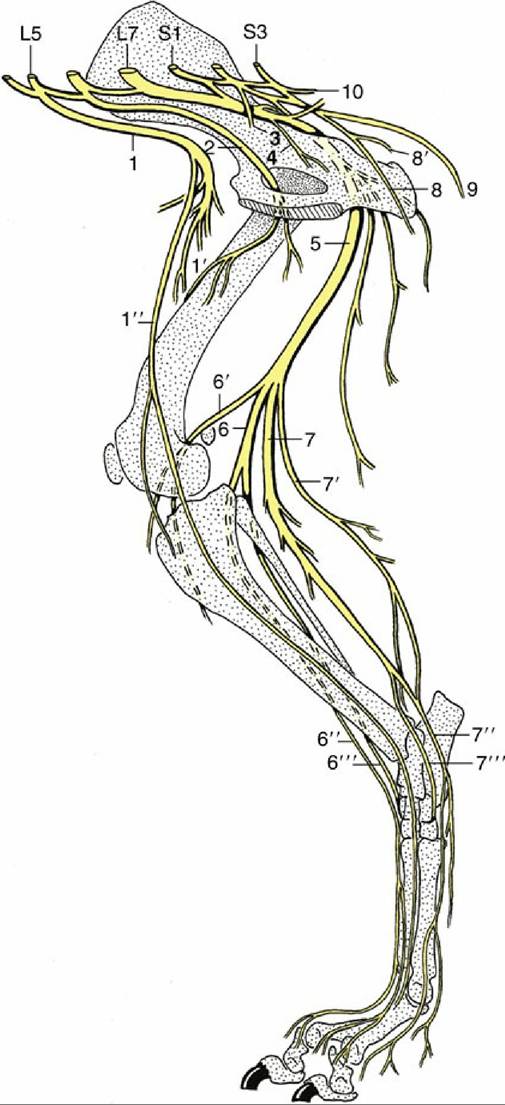
FIG. 8.75 The lumbar and sacral nerves of the dog; medial view. 1, Femoral n.; 1', branches to quadriceps; 1", saphenous n.; 2, obturator n.; 3, pelvic n.; 4, branch to obturator internus, gemelli, and quadratus femoris; 5, sciatic n.; 6, peroneal n.; 6', lateral cutaneous sural n.; 6", superficial peroneal n.; 6''', deep peroneal n.; 7, tibial n.; 7', caudal cutaneous sural n.; 7", medial plantar n.; 7''', lateral plantar n.; 8, pudendal n.; 8', deep perineal n.; 9, caudal cutaneous femoral n.; 10, caudal rectal n.; L5 and L7, fifth and seventh lumbar nn; S1 and S3, first and third sacral nn.
The remaining branches of the lumbosacral plexus arise from a common lumbosacral trunk that is largely formed by the last lumbar and first two sacral nerves, along with a smaller contribution from the penultimate lumbar nerve. The trunk leaves the pelvis through the greater sciatic foramen and almost at once detaches three branches before continuing on as the sciatic nerve.
The short cranial gluteal nerve supplies the tensor fasciae latae, the middle and deep gluteal, and in some species part of the superficial gluteal muscles, a group that—contrary to the usual expectation
—includes both flexor and extensor muscles of the hip.
The caudal gluteal nerve supplies the superficial gluteal muscle and the vertebral heads of origin of the hamstring muscles (biceps femoris, semitendinosus, and semimembranosus). These parts of the hamstrings are thought to represent an assimilation of elements of the superficial gluteal. It thus supplies extensor muscles of the hip.
The caudal cutaneous femoral nerve (Fig. 8.75/9) supplies skin over the caudal aspect of the thigh.
The sciatic nerve (Fig. 8.75/5) continues the lumbosacral trunk distally, passing between the middle and deep gluteal muscles before turning into the thigh caudal to the hip joint, where it is protected by the greater trochanter of the femur. It then runs between the biceps femoris laterally and the semitendinosus medially before dividing into its terminal branches, the common peroneal (fibular) and tibial nerves, at a level that varies among species. In the proximal part of its course, the sciatic detaches twigs to the internal obturator (unimportant except in ruminants and pigs), gemelli, and quadratus femoris (Fig. 8.75/4); other muscular branches that may appear to arise directly from the sciatic nerve are usually associated with its common peroneal and tibial divisions.
The common peroneal nerve (Fig. 8.75/6), the lesser of the terminal branches, arises from the lumbar roots of the lumbosacral trunk. It runs first with the tibial nerve but separates from this nerve to pass over the lateral head of the gastrocnemius to enter the leg. It detaches a branch, the lateral sural nerve (Fig. 8.75/6'), to the skin over the lateral aspect of the leg before dividing into superficial and deep branches when close to the head of the fibula. The superficial peroneal nerve (Fig. 8.75/6") supplies skin over the dorsal aspect of the leg and entire foot, except in the horse, in which it fades about the level of the fetlock joint. The deep peroneal nerve (Fig. 8.75/6''') supplies the dorsolateral muscles of the leg (flexors of the hock and extensors of the digits) and is also sensory to the structures of the foot. Because the sensory innervation of pedal structures varies considerably, the details are deferred to the accounts of individual species.
Paralysis of the common peroneal nerve produces overextension of the hock and flexion of the digits, which may be rested on their dorsal surfaces. The foot may be passively placed to support weight, and in time compensation may be possible (cf. radial paralysis, p. 310). There is also a considerable sensory deficit.
The tibial nerve (Fig. 8.75/7) arises from the sacral roots of the lumbosacral trunk. It detaches important proximal muscular branches to the pelvic heads of the hamstring muscles before freeing itself from the sciatic trunk to enter the leg by passing between the two heads of the gastrocnemius. About this level it first detaches a caudal sural nerve (Fig. 8.75/7') to the skin of this aspect of the leg and later detaches distal muscular branches to the gastrocnemius, soleus, popliteus, and caudal crural muscles. The nerve continues as an almost exclusively sensory trunk (although it will supply short digital muscles) within the fascial plate between the common calcanean tendon and the caudal crural muscles; it ends by dividing into medial and lateral plantar nerves when level with the point of the hock. The plantar nerves (Fig. 8.75/7" and 7''') continue into the plantar aspect of the foot to supply sensation to plantar structures chiefly but with some dorsal penetration that varies among species.
Section of or severe damage to the tibial nerve is manifest as overflexion of the hock and overextension of the digits. Similar damage to the sciatic trunk combines the effects of common peroneal and tibial nerve injuries, rendering the limb largely incapable, although fixation of the stifle joint by the unaffected quadriceps, supplied by the femoral nerve, may allow it to support some weight.
The Sacral and Caudal Ventral Rami
The sacral ventral rami caudal to and overlapping the roots of the lumbosacral plexus give rise to other important individual nerves. The pelvic nerves (Fig. 8.75/3), composed primarily of parasympathetic axons, are considered in the following section.
The pudendal nerve (Fig. 8.75/8) arises from various sacral nerves (S1-S3 in the dog, S2-S4 in ruminants, S[2]3-S4 in the horse). It is sensory to the rectum, perineal skin, and internal and external reproductive organs and motor to much of the striated perineal musculature. The nerve has both physiologic and applied importance, but because of species variation, it suffices here to say that the pudendal nerve takes an oblique course through the pelvis toward the ventral part of the pelvic outlet (see Fig. 29.5/7). The nerve provides deep and superficial perineal nerves in addition to various cutaneous branches and finally continues as the dorsal nerve of the penis (or clitoris). The superficial perineal branch supplies the skin of the anus, vulva, and ventral perineal region.
The deep perineal nerve supplies the ventral part of the striated musculature of the perineum, particularly that of the reproductive organs. The main trunk also supplies branches to the skin of the prepuce and scrotum in the male and of the caudal part of the udder in ungulates.
The caudal rectal nerves (Fig. 8.75/10) arise from the most caudal sacral nerves, sometimes overlapping the origin of the pudendal nerve. They supply sensory fibers to the rectum, anus, and perianal skin and motor fibers to the dorsal perineal striated musculature, including the levator ani. The division of territory between these nerves and the pudendal nerve is rather variable.
The ventral rami of the caudal nerves supply the ventral or depressor muscles of the tail.
In the standing animal, extensor muscles of the limbs must be continually active to support the body against gravity. Thus, damage to those nerves innervating the major extensor muscles of the limbs—that is, the radial, femoral, and sciatic nerves—produce obvious clinical signs and severe consequences for the animal.