TOPOGRAPHY
The brain and spinal cord are contained within a continuous space provided by the cranial cavity of the skull and the canal formed by successive bony rings and connecting ligaments and disks of the vertebral column.
The cranial cavity lies directly behind the nasal cavities. Smaller than is commonly supposed, its form and extent are not easily predicted from external inspection because the paranasal sinuses, horns, muscular ridges and other projections of the skull, and the temporal muscles all contribute significantly to the conformation of this part of the head. The closest agreement between the external contours and the cavity within the cranium is found in the newborn of all species; among adults this agreement is best retained in cats and in dogs of brachycephalic breeds. Fortunately, the exact location of the brain is rarely of practical significance except in the humane slaughter techniques mentioned in later chapters. It is probably sufficient to state meanwhile that the caudal limit of the cavity extends to the caudal wall of the skull—thickened by the frontal sinus in cattle— while the rostral limit shows considerable variation; it ends level with the caudal margin of the zygomatic processes of the frontal bones in dogs and cats and with the rostral level of these processes in horses and cattle, but it extends to the middle of the orbit in pigs and small ruminants.
The interior of the cranial cavity shows a fairly close correspondence with the contours of the brain, although significant intracranial space is required for the meninges and intermeningeal spaces that surround the brain and for the capacious intracranial venous sinuses. While the roof (calvaria) of the cavity remains largely undivided, the base is divided into three fossae; these need not be described in detail as the main features are depicted in Figure 8-54.
The rostral fossa is formed by the sphenoid and ethmoid bones and extends to the level of the optic canals, the passages of exit of the optic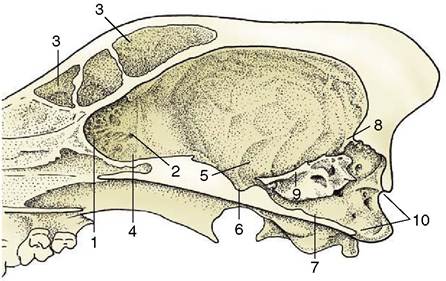
Figure 8-54 Sagittal section of the cranium of the dog. 1, Cribriform plate; 2, ethmoid foramen; 3, frontal sinus; 4, rostral fossa; 5, middle fossa; 6, hypophysial fossa; 7, caudal fossa; 8, tentorium cerebelli osseum; 9, petrosal crest; 10, foramen magnum.
nerves. It contains the olfactory bulbs, within recesses of the cribriform plate (Figure 8—54/7), and the rostral parts of the cerebral hemispheres. The middle fossa extends from the optic canals to the sharp petrosal crests (Figure 8-54/9) that project inward from the petrous temporal bones of the lateral walls. The floor is formed by the sphenoid bone, which carries the median hypophysial fossa (sella turcica) into which the hypophysis fits; it also presents various foramina of exit—the orbital fissure and the round and oval foramina—that were encountered in the previous description of the skull (p. 61). This, the widest part of the cranial cavity, contains the temporal and parietal lobes of the cerebral hemispheres. The caudal fossa extends from the caudal limit of the hypophysial fossa to the foramen magnum in the caudal wall. Its principal features are the contributions to its lateral walls made by the petrous parts of the temporal bones (each perforated by an internal acoustic meatus) and the jugular and hypoglossal foramina in the floor. The caudal fossa lodges the midbrain, pons, and medulla ventrally and the cerebellum dorsally.
The caudal, dorsal, and lateral walls of the entire cranial cavity blend together. Their most prominent internal feature is the tentorium cerebelli osseum (Figure 8-54∕the postmortem specimen, and by the internal vertebral venous plexus; the fat and vessels together cushion the spinal cord and allow it to adjust to the movements of the neck and back (see Figure 8-55).
The dural tube is attached at its caudal end, where the several meninges finally combine in a fibrous strand (filum terminale) that fuses with the upper surface of the caudal vertebrae.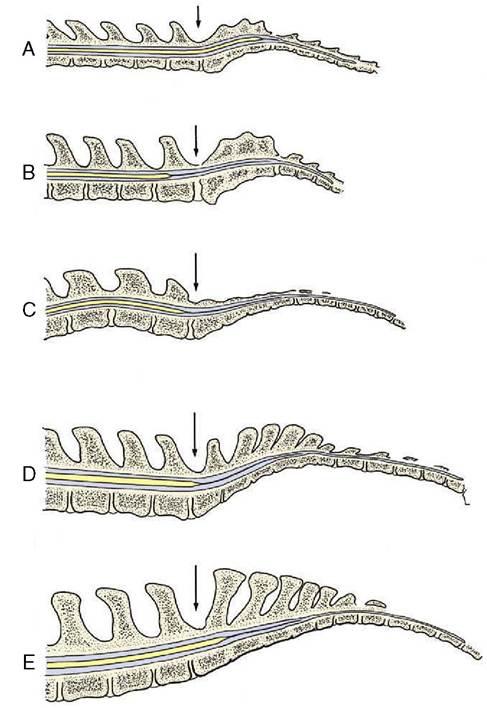
Figure 8-56 Median section of the vertebral canal and spinal cord of cat (A), dog (B), pig (C), cattle (D), and horse (E). The lumbosacral interarcuate space is indicated by an arrow. Notice the difference in caudal extent of the spinal cord in the different species. The thin extension of the spinal cord is the filum terminale that ends on the caudal vertebrae (not shown).
The fusion of the cranial dura with the periosteum obliterates the epidural space within the skull, and the cranial venous sinuses thus come to be enclosed within the thickness of the combined membrane. In addition to lining the cavity, the cranial dura forms certain folds that project inward and limit shuddering movements of the brain; these are a considerable hindrance to the removal of the intact brain at autopsy. One, the falx cerebri, extends from the dorsal and rostral cranial walls between the two cerebral hemispheres; caudally it joins a second, transverse fold, the membranous tentorium cerebelli, which separates the cerebellum from the cerebrum (Figure 8-57/7). The tentorium is ossified in its median part. A third specialization of the dura roofs the hypophysial fossa in which the hypophysis is seated, forming a diaphragm around the infundibular stalk.
A capillary space divides the dura from the subarachnoid, the first of the two more delicate inner membranes. This subdural space normally contains only a minute amount of a clear lymphlike fluid but may be enlarged by effusion of blood after an injury. The spinal part of the subdural space is crossed by a bilateral series of triangular (denticulate) ligaments that alternates with the origins of the spinal nerves; they attach the inner meninges to the dural tube and thus indirectly sling the cord (Figure 8-58/4).
The outer part of the arachnoid forms a continuous membrane molded against the dural envelope. Its inner surface is joined to the pia mater by numerous trabeculae and filaments, imaginatively compared to a spider’s web (the origin of the name subarachnoid). The pia mater is directly attached to the brain and cord and follows every change in their contours. The arachnoid space, which contains the clear, watery cerebrospinal fluid, is much wider than the subdural space but less uniform, particularly in its cranial part (see Figure 8-57).The widest parts (“cisterns”) of the cranial subarachnoid space are located between the more salient parts of the ventral surface of the brain and in the angle between the cerebellum and the dorsal aspect of the medulla. The dorsal widening, the cerebellomedullary cistern, is especially large and may be reached in the living animal if one passes a needle between the atlas and the skull (see Figure 8-57). Cisternal puncture is employed in both clinical and experimental work for obtaining samples of cerebrospinal fluid. The spinal subarachnoid space is more uniform but widens around the conus medullaris, which is a fortunate circumstance as access to the vertebral canal is easiest through the lumbosacral interarcuate space (Figure 8-59).
The pia mater is firmly attached to the outer surface of the brain and cord, and many branches from arteries within the pia penetrate the brain and cord substance. These vessels are initially enclosed by pial sleeves but these soon merge with the vascular walls. A thickening of the pia fills the ventral fissure of the spinal cord, where it appears as a glittering silver line.
All three meninges form cuffs around the roots of origin of the cranial and spinal nerves.
The cerebrospinal fluid within the arachnoid space forms a water jacket that buoys up and protects the soft brain and cord. It is largely a product of the ependymal lining of the ventricular system within the brain, and the overwhelmingly larger part is produced where this covers the choroid plexuses, vascular tufts that invaginate the ventricles (Figure 8-60/6,9).
An additional contribution to the fluid is made by the pial vessels.The ventricles are local modifications of the lumen of the neural tube; they have complicated shapes, but as these are illustrated (Figure 8-61) and as the details have little veterinary significance, they need not be
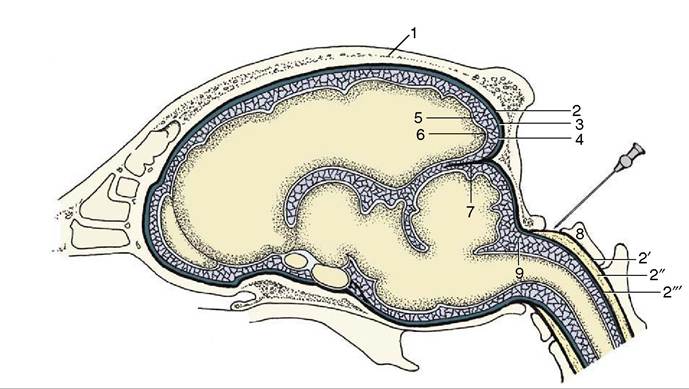
Figure 8-57 Schematic representation of the meninges of the brain. The needle points to the atlantooccipital space and the cerebellomedullary cistern. 1, Calvaria; 2, dura mater (also connected to the bone as periosteum); 2', periosteum of vertebral canal; 2'', epidural space (with fat); 2'", dura mater of spinal cord; 3, subdural space; 4, arachnoid; 5, arachnoid space; 6, pia mater; 7, membranous tentorium cerebelli; 8, atlas; 9, cerebellomedullary cistern.
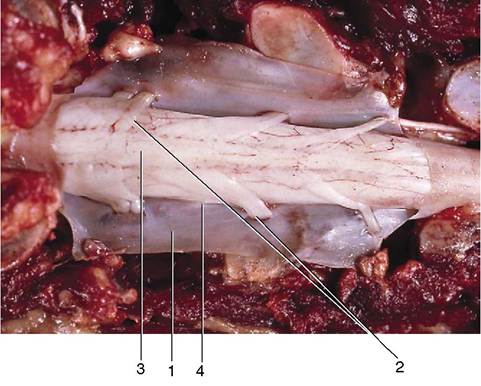
Figure 8-58 Dorsal view of the opened vertebral canal. The dura mater has been dissected and is reflected. 1, Dura mater; 2, dorsal rootlets of a spinal nerve; 3, spinal cord (covered by pia mater); 4, denticulate ligament.
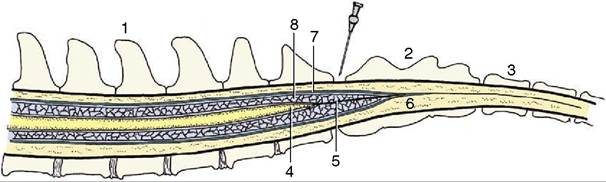
Figure 8-59 Schematic median section of the vertebral canal and its contents. The needle points to the lumbosacral interarcu- ate space. 1, Lumbar vertebra; 2, sacrum; 3, caudal vertebra; 4, conus medullaris; 5, filum terminale; 6, epidural space; 7, dura mater; 8, arachnoid space with cerebrospinal fluid.
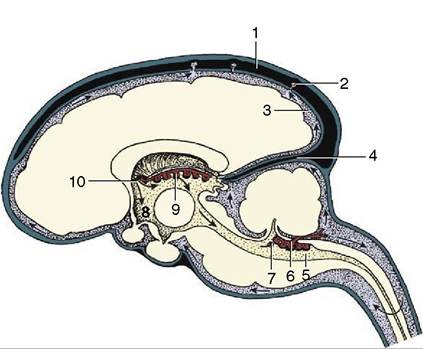
Figure 8-60 The production and circulation of cerebrospinal fluid (sagittal section). The blood vessels are in black, the subarachnoid spaces are heavily shaded, the ventricles are lightly shaded, and the nervous tissue is white. The direction of the flow of the cerebrospinal fluid is indicated by arrows.
The cerebrospinal fluid is secreted by the choroid plexus (6,9) of the lateral, third, and fourth ventricles. It escapes into the subarachnoid space via the aperture of the fourth ventricle (7). The cerebrospinal fluid is transferred to the systemic circulation (1) at the arachnoid villi (2). 1, Dorsal sagittal sinus; 2, arachnoid villus; 3, subarachnoid space; 4, membranous tentorium cerebelli; 5, fourth ventricle; 6, choroid plexus of fourth ventricle; 7, aperture of fourth ventricle; 8, third ventricle; 9, choroid plexus of third ventricle; 10, interventricular foramen, connecting the lateral and third ventricles
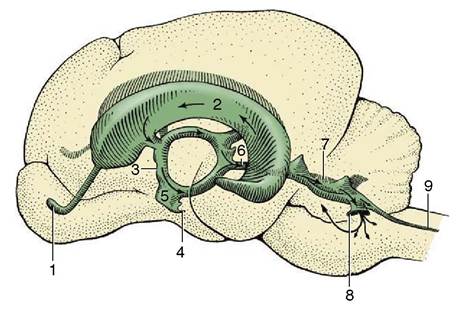
Figure 8-61 Lateral view of a cast of the ventricles of the brain of the dog. 1, Cavity of olfactory bulb; 2, lateral ventricle; 3, third ventricle; 4, infundibular recess; 5, optic recess; 6, mesencephalic aqueduct; 7, fourth ventricle; 8, lateral recess; 9, central canal.
described. It is more important to understand their relationship to the choroid plexuses. The plexuses of the lateral and third ventricles, which merge within the interventricular foramen, develop within a fold of the pia that becomes entrapped between the expanding tel-
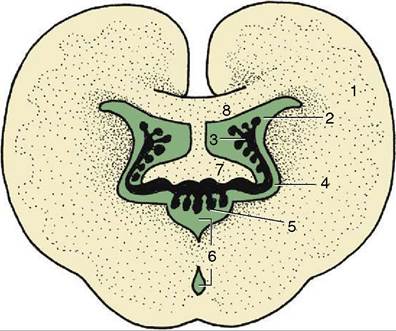
Figure 8-62 Schematic section of the brain illustrating the interrelations of the third and lateral ventricles and their choroid plexuses. 1, Cerebral hemisphere; 2, lateral ventricle; 3, choroid plexus of lateral ventricle; 4, interventricular foramen; 5, choroid plexus of third ventricle; 6, third ventricle; 7, fornix; 8, corpus callosum.
encephalic vesicles and the roof of the diencephalon (Figure 8-62). The plexuses of the fourth ventricle develop separately within the pia over the caudal medullary velum. In the course of development these plexuses thrust themselves into the lumen of the fourth ventricle; parts later reemerge into the arachnoid space by herniating through paired lateral openings in the roof (Figure 8-63).
The clear colorless cerebrospinal fluid is to a certain extent also formed from the blood plasma by ultrafiltration through the “blood-cerebrospinal fluid barrier” (blood-brain barrier) composed of vascular endothelial cells. The fluid has a higher concentration of potassium and calcium ions and a lower concentration of sodium, magnesium, and chloride ions than the plasma; it is also rather deficient in glucose and, most importantly, contains little protein because the barrier is impermeable to larger molecules, which of course include those of many antibiotic and other drugs.
In addition to its mechanical role, the cerebrospinal fluid protects the brain through its chemical buffering capacity, which provides a rather stable milieu. It also transports nutrients, flushes away waste products, and serves as a medium for the diffusion of neuroendocrine and neurotransmitter substances.
The fluid is produced continuously, at a rate of some 30 mL per hour in the dog, and first circulates through the ventricular system, moved onward by the filtration pressure and ciliary activity of the ependymal lining. It then escapes from the interior of the brain through the lateral apertures of the fourth ventricle (Figure 8-60/7; in some species there is a third median opening unre-
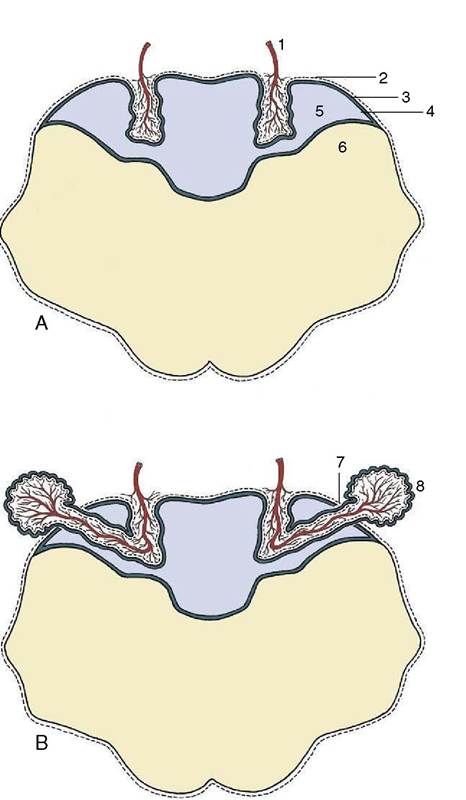
Figure 8-63 The formation of the choroid plexus in the roof of the fourth ventricle (A) and its later extension into the subarachnoid space (B). 1, Blood vessel invagination; 2, pia mater; 3, caudal medullary velum; 4, ependyma; 5, fourth ventricle; 6, myelencephalon; 7, aperture of fourth ventricle; 8, choroid plexus extending into subarachnoid space.
lated to the plexus). The fluid bathes the brain and cord before returning to the blood, mostly through the arachnoid granulations (villi) (Figure 8-64/70), projections of the arachnoid and subarachnoid space that pierce the dura to enter the dorsal sagittal venous sinus of the brain; these formations become increasingly prominent with age. (Obliteration of the villi results in hydrocephalus because drainage of the fluid is hampered while its production continues and is not influenced by a feedback mechanism.) A smaller part of the fluid percolates along the meningeal cuffs that surround the cranial and spinal nerves at their origins and is eventually absorbed by perineural lymphatics; these connections are believed to provide potential routes for the retrograde (i.e.,
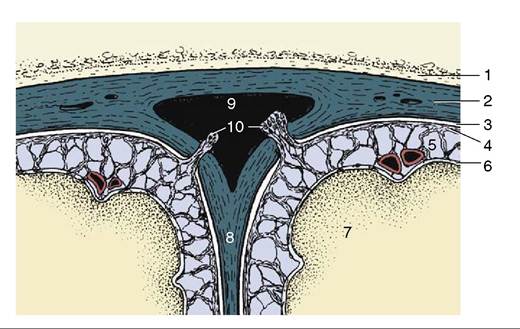
Figure 8-64 Transverse section of the dorsal sagittal sinus and adjacent meninges. Cerebrospinal fluid is transferred from the subarachnoid space to the sinus via the arachnoid granulations (villi). 1, Roof of cranial cavity; 2, fused dura mater and periosteum; 3, subdural space; 4, arachnoid; 5, subarachnoid space; 6, pia mater; 7, cerebral hemisphere; 8, falx cerebri; 9, dorsal sagittal sinus; 10, arachnoid granulations (villi).
toward the meninges and nervous tissue) spread of infection.