THE ARTERIAL BLOOD SUPPLY
The blood supply to the brain comes mainly from the circulus arteriosus cerebri (formerly known as the circle of Willis), which lies ventral to the hypothalamus where it forms a ring around—but at some distance from—the infundibular stalk.
The appearance of the circle and the pattern of its major branches are remarkably constant among mammals, although the sources from which the circle is supplied and the directions in which blood flows in certain vessels vary. For this reason, the initial account is based on the arrangements in the dog, an animal in which the arrangement is not only relatively simple but also of the most common pattern.The arterial circle of the dog is supplied from three sources: paired internal carotid arteries laterally and the basilar artery caudally (Figure 8-65). The internal carotid artery (Figure 8-65/5) is a terminal branch of the common carotid from which it springs opposite the pharynx. It then runs toward the base of the skull. In many species the artery makes immediate entry to the cranial cavity through a carotid foramen in the cranial floor, but in the dog it must first traverse a tunnel (carotid canal) in the bone medial to the tympanic bulla. The artery is released at the rostral end of the tunnel and describes a loop that first carries it ventrally, then dorsally, before it finally gains the cranial cavity. It then penetrates the outer meninges, which involves passage through the cavernous venous sinus enclosed within a splitting of the dura, before dividing into divergent
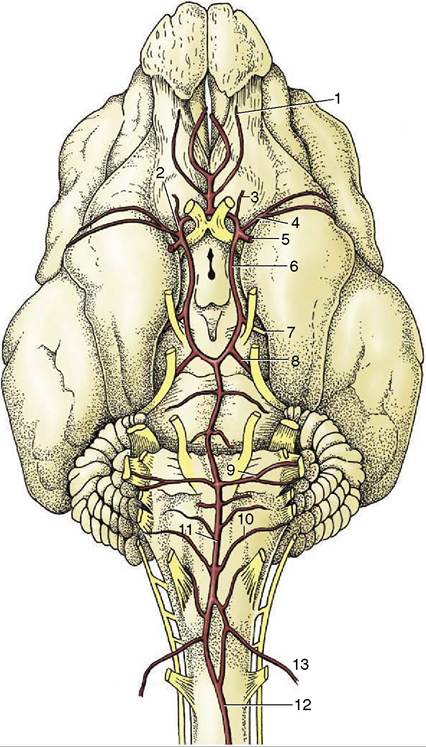
Figure 8-65 Arteries on the ventral surface of the canine brain. 1, Internal ethmoidal a.; 2, rostral cerebral a.; 3, internal ophthalmic a., 4, middle cerebral a.; 5, internal carotid a.; 6, caudal communicating a.; 7, caudal cerebral a.; 8, rostral cerebellar a.; 9, labyrinthine a.; 10, caudal cerebellar a; 11, basilar a.; 12, ventral spinal a.; 13, vertebral a.
branches. The rostral branch unites with its fellow to complete the rostral half of the circle, the half from which the large rostral and middle cerebral arteries arise. The caudal branch anastomoses with a branch of the basilar artery (which reaches the circle along the midventral surface of the brainstem) to complete the circle (Figure 8—65/11). The caudal cerebral and rostral cerebellar arteries leave the caudal half of the circle; the fifth major artery to the brain, the caudal cerebellar, leaves the basilar artery directly.
The blood within the basilar artery has a composite origin. The artery appears to be the direct continuation of the small ventral spinal artery but is greatly reinforced by anastomosis with the vertebral artery (Figure 8—65/13), which passes into the vertebral canal through the atlas. The vertebral artery itself receives anastomotic branches (dog and horse) from the occipital artery (another branch of the common carotid) before entering the canal, and it would thus appear that this vessel (occipital artery) also contributes to the supply of the brain. However, the vertebral artery is the main if not sole supply to the occipital lobes of the cerebral hemispheres and other caudal parts of the brain.
The arrangement is more complicated in many other species. In these the internal carotid connects with other arteries of the head, especially the maxillary, before discharging into the arterial circle. The anastomosis may be small initially, but in many species it later enlarges and detaches many tortuous branches, which together substitute for the original single channel. This arrangement, which may present a rather tangled appearance, is known as a rete mirabile and has a rather enigmatic significance; the arrangement enhances the efficiency of the bloodcooling mechanism that is discussed shortly. In some species the lumen of the part of the internal carotid artery proximal to the rete becomes obliterated, sometimes only a considerable time after birth; when this happens, the emissary artery from the rete delivers blood that is wholly of external carotid origin (see Figure 7-35).
This arrangement is found in both sheep and cattle, although these species differ in other features of the arterial supply to the brain (p. 661).The brain, particularly its gray substance, has very high metabolic requirements, and the arterial supply is commensurate with this, amounting to 15% or 20% of the cardiac output. Despite this, the vessels that actually penetrate the brain are uniformly small, which is a feature that may be related to the need to avoid large, pulsating trunks within the delicate brain tissue. Moreover, in sharp contrast to the wide anastomoses between the feeding vessels, any intracerebral anastomoses are narrow and mostly connect functional end-arteries. This fact, coupled with the very limited regeneration capacity of brain tissue, explains why the most serious consequences may attend occlusion or rupture of a small vessel that may be the sole effective supply to some vital nucleus or tract. Notorious examples are provided by the small arteries within the human corpus striatum, where an infarct is so often the cause of a stroke.
The permeability of the blood capillaries of the nervous tissue is reduced, resulting in the blood-brain barrier. The main structural components of this barrier are provided by the continuity between the endothelial cells of these capillaries and pericytes, astrocytes, and the basement membrane surrounding these capillaries (see p. 217; Figure 8-66).
The spinal cord is supplied by three arteries that run its length. The largest, the ventral spinal artery, follows the surface of the ventral fissure of the cord; paired
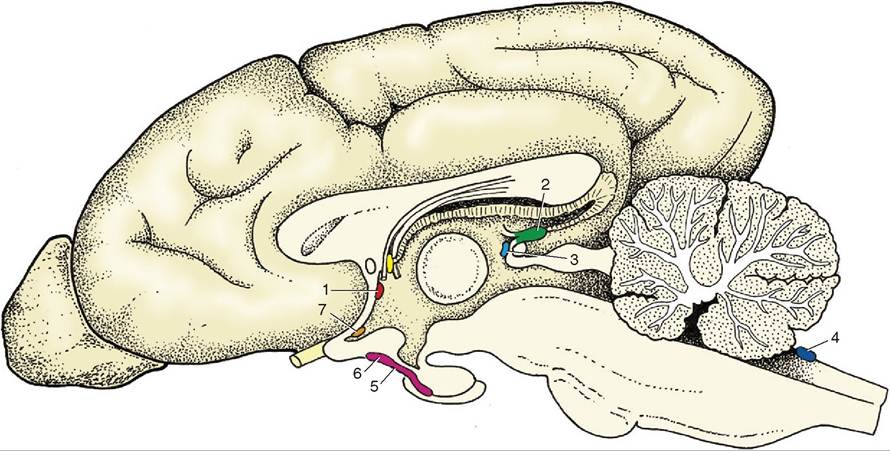
Figure 8-66 Schematic median section of the canine brain with an indication of the locations of the Circumventricular organs. 1, Subfornical organ; 2, pineal body; 3, subcommissural organ; 4, area postrema; 5, posterior and intermediate lobes of pituitary; 6, median eminence; 7, vascular organ of lamina terminalis.
dorsolateral spinal arteries run close to the furrow from which the dorsal roots of the spinal nerves arise. All three are periodically reinforced by branches from regional arteries: vertebral in the neck and intercostal, lumbar, and sacral in the trunk. These enter at the intervertebral foramina, often in the form of narrow vessels that accompany the roots of the spinal nerves; they form plexuses on the surface of the cord with which the major longitudinal arteries connect. This theoretically regular pattern is subject to much variation, both specific and individual, in which many expected reinforcing arteries are lacking, the plexus is unevenly developed, and stretches of the longitudinal trunks are attenuated.
Branches of the ventral spinal artery supply the “core” of the cord, the gray substance, and the adjacent layer of white substance by an approach through the ventral fissure (see Figure 19-5). The greater part of the white substance is supplied by radial twigs from the dorsolateral arteries and surface plexus. Internal anastomoses between the two sets of vessels, although common, are of questionable efficiency.